Реферат: Функциональная архитектура зрительной коры
Функциональная архитектура зрительной коры
Функциональная архитектура зрительной коры
Зрительная кора организована в виде вертикальных кластеров клеток, имеющих сходные функциональные свойства. Нейроны, получающие информацию избирательно либо от правого, либо от левого глаза, сгруппированы в глазодоминантные колонки. Ориентационные колонки состоят из нейронов, которые имеют сходную чувствительность к углам ориентации линий и краев изображения. Глазодоминантные колонки и ориентационные колонки были впервые открыты при регистрации электрической активности от группы кортикальных клеток во время прохождения электродом вглубь зрительной коры. Глазодоминантные и ориентационные колонки могут быть также визуализованы при помощи биохимических и оптических методов, которые выявляют зоны активности коры у живого животного.
Аксоны крупноклеточных (magnocellular. M) и мелкоклеточных (parvocellular, P) нейронов ядер латерального коленчатого тела проецируются в различные области 4 слоя первичной зрительной коры. В дальнейшем эта информация по М- или Р-трактам распространяется в различающиеся области первичной и вторичной зрительной коры. Нейроны М-пути отвечают за детекцию движущихся стимулов. Они также чувствительны к различиям в контрастности и интенсивности зрительных стимулов. Нейроны Р-пути ответственны за тонкие детали изображения и восприятие цвета.
При восприятии изображения такие его элементы, как цвет и движение, анализируются независимо. Это иллюстрируется тем фактом, что повреждения в изолированных зонах мозга приводят к селективной потере одного из этих свойств, но не сказываются на общем восприятии изображения глазом. Повреждения в области париетальной коры, известной как МТ (или V5), приводят к полной потере способности определять движения объектов и к нарушениям в восприятии интенсивности изображения. При локализации повреждения в затылочно-височной области (зона V4) теряется способность к распознаванию цвета.
Выдающимся достижением является использование неинвазивных методов функционального магнитного резонанса для определения зон активности мозга животных и человека. Хотя отдельные корковые глазодоминантные и ориентационные колонки имеют размеры меньше сегодняшнего предела разрешения данного метода, именно при помощи этого метода были найдены области зрительной коры, специализирующиеся на задачах особого рода, таких как определение движения и распознавание лиц.
Путем анализа эффекта зрительных стимулов на последовательно связанные кортикальные клетки можно понять процессы переработки информации в первичной зрительной коре. Этот подход приблизил нас к пониманию клеточных механизмов анализа формы в каждой отдельной точке поля зрения. Сейчас же нашей задачей является изучить, каким образом в коре кодируются другие аспекты восприятия, такие как цвет и движение, и каким образом эти свойства могут быть затем собраны воедино при восприятии цельного изображения.
Мы уже описали поточечное представление сетчатки в области зрительной зоны (V1) и то, каким образом происходит разделение информации от обоих глаз в колонках глазного доминирования. В пределах данной ретинотопической карты существует функциональное деление; так, например, определенные колонки клеток реагируют на линии строго определенной ориентации. Мы начинаем эту главу исследованием взаимосвязи между колонками глазного доминирования и ориентационными колонками, затем мы рассмотрим доказательства того, что в зрительной коре движение и цвет анализируются параллельно различными группами клеток. И, наконец, мы рассмотрим примеры высшего уровня обработки зрительной информации в зонах, расположенных вне первичной зрительной коры.
Колонки с доминированием одного глаза и ориентационные колонки
В ранних экспериментах Хьюбеля и Визеля было показано, что клетки коры со сходными свойствами образуют между собой связи, формируя вертикально организованные колонки. В экспериментах с проникающим электродом, при его продвижении вглубь зрительной коры, у всех клеток обнаруживалась одна и та же ориентация оси рецептивного поля, то же глазное доминирование и одно и то же расположение рецептивного поля. Мы уже упоминали колонки предпочтения определенного глаза (eye preference).Сигналы от обоих глаз разделяются в пределах слоя 4, где кортикальные нейроны получают сигналы только от одного глаза. В любой данной колонке, выходящей выше или ниже слоя 4, все кортикальные нейроны, даже те, которые получают информацию от обоих глаз, имеют одинаковое глазное предпочтение (либо левый, либо правый глаз). Таким образом, существуют колонки с предпочтительной, или по-другому, с доминирующей реакцией на сигналы от одного определенного глаза (глазодоминантные колонки). На рис.1 иллюстрируются различия в таком глазном доминировании в нейронах стриарной коры обезьяны. Клетки (всего 1116) подразделяются на 7 групп. Группы 1 и 7 получают информацию только от одного из глаз и находятся в слое 4 коры. В группах 2, 3, 5 и 6 эффект одного из глаз сильнее, нежели другого, и только в средней группе 4 оба глаза имеют примерно одинаково выраженные эффекты. Из гистограммы также ясно, что большинство клеток отвечают преимущественно на сигналы от правого или левого глаза.

Рис. 1. Физиологическая демонстрация глазодоминантных колонок.
Глазодоминантные колонки формируют своеобразный перемежающийся рисунок зрительной коры. Паттерны глазного предпочтения в коре могут быть непосредственно визуализованы, используя оптические сигналы от искусственно введенных красителей. При помощи этого метода можно исследовать активность больших участках коры во время предъявления животному различных зрительных стимулов. На рис. 2 показаны глазодоминантные колонки, выявленные в таком эксперименте. Полосатый (перемежающийся) рисунок активности напоминает тот, который может быть получен при введении радиоактивных меток в один глаз . При предъявлении зрительного стимула только одному глазу выявляются полоски клеток, получающих сигналы только от этого глаза, отделенные друг от друга клетками с невысоким уровнем активности. Проекции этих полосок от поверхности коры вглубь образуют колонки глазного доминирования, делящие ретинотопическую карту на более мелкие области. Таким образом, информация о предмете, находящемся в одном и том же месте зрительного поля, идущая от обоих глаз, интегрируется в пределах зрительной коры как соседними клетками, так и соседними колонками (рис. 1В).
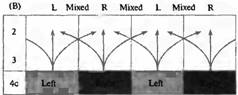
(А) Глазные предпочтения е 1116 клетках в зоне V1 у 28 макак резус. Большинство клеток (группы со второй по шестую) получают информацию от обоих глаз. (В) Диаграмма, показывающая каким образом информация от двух глаз, направляющаяся в слой 4 коры, комбинируется в более поверхностных слоях при помощи горизонтальных и косых соединений, дающих клетки с бинокулярными полями.
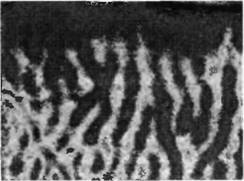
Рис. 2. Выявление глазодоминантных колонок с помощью оптической регистрации. Чувствительная камера обнаруживает оптический сигнал в коре обезьяны во время стимуляции только одного глаза. Изменения в интенсивности сигнала кодируются при помощи цвета, поэтому активные зоны выглядят в виде светлых полос. Паттерн светлых полос соответствует колонкам глазного доминирования, выявляемым при маркировании анатомическими методами.
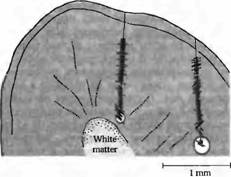
Рис. 3. Оси ориентации рецептивных полей нейронов по мере погружения электрода вглубь коры мозга кошки. Все последующие клетки демонстрируют одинаковую ориентацию оси, что показано в виде угла, под которым располагается штриховая полоска по отношению к линии проникновения электрода. Погружение электрода справа идет с большим отклонением от перпендикуляра, следовательно, при пересечении им нескольких колонок часто меняется и ориентация оси. Расположение каждой клетки определяется при помощи нанесения небольших повреждений по мере погружения электрода и при завершении погружения (показано кружком). На основании этих данных производится реконструкция хода электрода. Подобные эксперименты позволили установить, что клетки со сходными свойствами у кошек и обезьян организованы в виде колонок, идущих под определенными углами к поверхности коры.
Ориентационные колонки
Какие другие функциональные объединения возможны для зрительных нейронов в зоне V1?
Ориентационные предпочтения простых и комплексных клеток. Таким образом, логичен вопрос: является ли данная особенность систематически повторяющейся в зрительной коре? Репрезентативный эксперимент, отвечающий на этот вопрос, представлен на рис. 3. Микроэлектрод погружается перпендикулярно поверхности вглубь серого вещества в зоне V1 мозга кошки. Каждый штрих на этом рисунке показывает расположение одной клетки и предпочтительную ориентацию ее рецептивного поля при предъявлении визуальных раздражителей в виде полосок. После погружения электрода в контрольной точке клетки локально повреждаются посредством пропускания электрического тока. Расположение каждой регистрируемой клетки затем реконструируется на основании этих контрольных точек (конечная точка показана кружком в конце пути электрода). Как показано с левой стороны рисунка, первые (начиная с поверхности) 38 клеток наиболее эффективно активировались полосками, расположенными под углом около 90 градусов к вертикальной оси. После погружения на глубину около 0,6 мм ось ориентации рецептивного поля клеток сместилась примерно на 45 градусов. При другом погружении, показанном справа, с более острым углом к поверхности коры, каждая последующая клетка имеет разные варианты ориентации оси. При таком наклонном проникновении оси полей меняются по мере прохождения через разные колонки, имеющие отличающуюся ориентацию. Ориентационные колонки получают сигналы от клеток, рецептивные поля которых значительно перекрываются на поверхности сетчатки.
Информация об организации ориентационных колонок в зрительной коре обезьян и кошек была впервые получена при проведении именно косых (тангенциальных), а не перпендикулярных погружений электрода в глубь коры. При продвижении электрода на каждые 50 мкм в горизонтальном направлении вдоль коры ориентация оси рецептивного поля меняется примерно на 10 градусов, иногда такие последовательные смены ориентации достигают в конечном итоге 180 градусов. Колонки ориентации (20-50 мкм) гораздо уже, чем глазодоминантные колонки (250-500 мкм). Первая анатомическая демонстрация ориентационных колонок, основанная на их разной функциональной активности, была произведена Соколовым, который использовал 2-деоксиглюкозу, хорошо захватывающуюся активными клетками. В результате метаболически активные клетки становились радиоактивными и их расположение можно было видеть на ауторадиограммах. У обезьян и кошек, которым показывали горизонтальные или вертикальные полосы, «полоски» радиоактивности совпадали с расположением колонок вертикальной или горизонтальной ориентации, различимых в коре без всякой окраски. Организация ориентационных колонок изучалась также при помощи оптических методов на живых животных. Примером может служить эксперимент Бонхоеффера и Гринвальда. Представление зрительных стимулов различной ориентации приводило к активности в различных кортикальных зонах. Ответы на разные ориентации показаны разными цветами. Удивительной является организация ориентационных колонок по отношению друг к другу. Сначала их организация кажется достаточно беспорядочной. Однако при тщательном исследовании можно заметить определенные центры в виде пестрого «волчка», в котором сходятся отдельные точки всех ориентаций. Отсюда клетки, ответственные за определенную ориентацию, расходятся в удивительно правильном порядке. Некоторые волчки ориентированы по часовой стрелке, некоторые — против. Таким образом, ориентация в коре представлена скорее в радиальном, чем в линейном виде. Каждый угол ориентации уникален (возникает только один раз) в пределах цикла, вертикальные и горизонтальные контуры занимают несколько большую площадь коры. На каждом квадратном миллиметре коры находятся один или два таких центра, равноудаленных друг от друга. Интересно то, что подобный способ организации коры был предложен ранее, на основании теоретических предпосылок.
Связь между колонками глазного доминирования и ориентационными колонками
При помощи методов оптической микроскопии была также выявлена взаимосвязь между ориентационными колонками и колонками глазного доминирования. Структура корковой активности регистрировалась сначала при стимуляции одного глаза, а затем определялась снова при помощи серии ориентированных световых полос. Результаты одного такого эксперимента .Каждый контур, имеющий свою ориентацию, показан в виде отдельной цветной линии (и изоориентированного контура), зоны глазного доминирования показаны в виде светлых или темных участков. Волчки ориентации четко видны в виде сходящихся изоориентированных контуров, при этом несколько контурных линий между различными волчками обычно пересекают границу глазного доминирования. Таким образом, получается, что большинство ориентационных зон оказывается расщеплено на контралатеральную и ипсилатеральную половинки, получая информацию от обоих глаз с данного места зрительного поля. На самом деле, центр каждого волчка стремится оказаться наиболее близко к центру глазного доминирования, а контуры изоориентации имеют свойство пересекать границы глазного доминирования под определенными углами .
Параллельная обработка информации о форме, движении и цвете
Колонки зрительного доминирования и ориентационные колонки представляют собой только два типа возможной функциональной организации нейронов зрительной коры. Направление движения, пространственная частота (в основном зависящая от размеров рецептивного поля), несоразмерность изображения (важная детерминанта для восприятия глубины изображения) также обеспечиваются функциональными образованиями зрительной коры, имеющими вид колонок. Возникает логичный вопрос: каким образом выполняются все необходимые виды анализа изображения для каждой точки в зоне коры, куда проецируется ретинотопическая карта? Эффект «смешивания» функциональных колонок, дает частичный ответ на этот вопрос. В самом деле, задолго до использования оптических методов для поиска подобных взаимосвязей, Хьюбель и Визель предложили концептуальную схему того, что они назвали «сверхколонка» (hypercolumn). Под этим термином они понимали такое объединение нейронов, в котором представлены все возможные ориентации соответствующих областей зрительных полей обоих глаз. Согласно этому представлению соседние сверхколонки анализируют информацию таким же образом, как и соседние перекрывающиеся части зрительных полей. Тем не менее остается пока трудной задачей объединить эту точку зрения с данными современных методов анализа изображения.
Проекции в зрительную зону 2 (V2)
При окрашивании на цитохромоксидазу зоны V2 выявляется паттерн, отличный от того, который виден в зоне V. Окрашивание происходит в виде серии тонких и толстых полосок, перемежающихся с более бледными зонами с меньшей активностью фермента (рис. 4. Эти параллельные полоски идут под определенными углами от границы между зонами V1 и V2. После введения фермента пероксидазы хрена в окрашенные зоны V1, он захватывается терминалами аксона и транспортируется ретроградно, показывая, что нейрон, отростки которого проецируются в пределах зоны «пятна», расположен в пределах тонкой полосы окрашивания зоны V2 зрительной коры. Эти связи взаимны: при инъекции в тонкие полоски возникает окрашивание в области «пятен». И наоборот, области между «пятнами» образуют связи с бледными широкими полосами, которые получают в основном информацию от больших клеток из слоев 4В и 4С. Примечательно, что это функциональное подразделение может быть выявлено даже на молекулярном уровне: моноклоналъные антитела Cat-301 в основном маркируют крупноклеточные пути в пределах зрительной коры обезьяны.

Рис. 4. «Пятна» в зоне V1 и «полоски» в зоне V2 зрительной коры обезьян, окрашенной на цитохромоксидазу. Пятна организованы в виде узора «в горошек». Можно различить четкую границу между зонами V1 и V2 . По этой линии «пятна» переходят в «полоски», толстые и тонкие, идущие под определенным углом к границе.
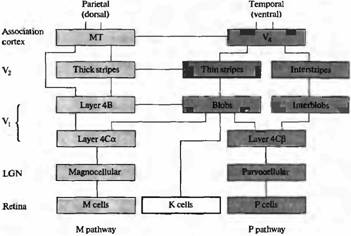
Рис. 5. Схематическая организация М-, Р- и К-кана-лов, направляющихся в зрительную кору. Функционально различные слои ЛКТ направляют отростки к различным слоям в V1. К-слои проецируются в «пятна» в слое 2 и 3. М- и Р-слои области 4С могут взаимодействовать предпочтительно с областями «пятна» и с областями, расположенными между пятен в слоях 2 и 3. «Пятна» проецируются преимущественно в тонкие полоски в V2 . Тонкие полоски проецируются в V4. Толстые полосы в V2 получают информацию от слоя 4В в V и проецируются в ассоциативную зону МТ (V5). M-каналы проецируются в дорзальную (теменную) зрительную кору, где происходит анализ движения. Р-каналы проецируются предпочтительно в зону V4, где осуществляется обработка информации о цвете.
Ассоциативные зоны зрительной коры
Значительные усилия были потрачены для понимания процесса переработки информации в М- и Р-каналах зрительной коры. Подобные исследования были вызваны функциональными и анатомическими различиями ганглиозных клеток сетчатки и нейронов коленчатого тела (рис. 4). Дополнительная информация обеспечивалась психофизическими экспериментами, исследованием строения головного мозга, изучением пациентов с различными мозговыми повреждениями. Все это привело к предположению, что различные типы анализа зрительной информации производятся в физически изолированных регионах мозга, в так называемых ассоциативных зонах коры. Дорзальный или париетальный тракт (называемый так потому, что соответствующие зоны коры находятся в теменной доле, расположенной более дорзально по отношению к первичной зрительной коре), как считают, является важным звеном в оценке движения и пространственного взаиморасположения форм — свойств, сходных со свойствами М-каналов, которые мы уже описали. Повреждения дорзального, париетального тракта выражаются в игнорировании участка зрительного поля и нарушении видеомоторной ориентации. Повреждения в области височной коры (вентральнее V1) уменьшают способность к зрительной идентификации объектов, их цветов и тонких деталей — отражая свойства Р-каналов.
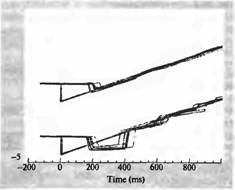
Рис. 6. Организация зрительного движения в зоне МТ Обезьяна была обучена отслеживать взглядом движущиеся предметы (движение стимула показано светлой серой линией). На верхней записи показано положение глаз обезьяны относительно движущегося объекта. После начального резкого быстрого движения предмет проецируется на зрительной ямке (быстрое отклонение глаза вниз), затем глаз точно следует за движением предмета. Однако после введение нейротоксина в область МТ (нижняя запись) первое движение является избыточным и «проскакивает» цель. Дальнейшее слежение идет медленнее, чем это необходимо, как будто была неправильна рассчитана скорость движения предмета.
Детекция движения и зона МТ
Если попросить дать краткое определение, что такое зрение, большинство из нас скажет о восприятии объектов, включающем анализ тонких деталей, примерно как это происходит во время чтения этих слов. Однако не менее важной функцией зрения является анализ движения, хотя эта функция и остается в основном непроизвольной и подсознательной (за исключением тех, кто из-за окуломоторного расстройства не может автоматически управлять перемещением зрительного поля). Как уже было описано, движение анализируется крупноклеточно-париетальной частью зрительного тракта (рис. 4). Нейроны крупноклеточного пути чувствительны к движущимся стимулам, это их свойство сохраняется в зонах V1 и V2 (в пределах толстых полос), через которые этот путь проходит, направляясь к средневисочной ассоциативной коре (зона МТ, или V5). Зона МТ также имеет ретинотопическое строение. Нейроны в этой области чувствительны к скорости и направлению движения стимула, а также объединены в группы в виде колонок со сходными предпочтительными направлениями движения. Когда небольшие области МТ химически повреждаются при помощи нейротоксина, у обезьяны нарушается способность воспринимать характер движения точек в соответствующих областях зрительного поля, хотя чувствительность к контрастности изображения при этом не меняется.
Зона МТ и зрительное слежение
Зона МТ играет важную роль в процессе зрительного слежения. Это было показано рядом экспериментов, в которых обезьяну обучали следить глазами за движущимся предметом (рис. 5). Нормальный паттерн движения глаз показан на верхней записи на рис. 5. При этом при движении объекта (траектория начинается от точки 0) глаза фокусировались на нем при помощи быстрого, отрывистого движения (с отклонением вниз через 200 миллисекунд), а затем при помощи аккуратного слежения предмет удерживался в области зрительной ямки. После введения небольшого количества нейротоксина (иботеновой кислоты) в зону МТ, в область представительства центральной ямки, способность обезьяны следить за движущимся объектом значительно нарушалась. В частности, после первого быстрого движения дальнейшая скорость слежения была значительно ниже скорости перемещения объекта. Нарушено было также и само начальное установочное движение глаз (рис.5, нижний пробег). Таким образом, повреждение зоны МТ приводило к тому, что животное теряло способность адекватно отслеживать движение объекта.
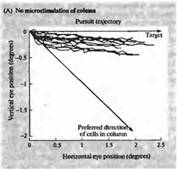
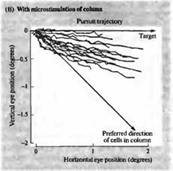
Рис. 6. Изменение направления движения глаза при электрической стимуляции в области МТ (А) Запись движения глаза в ответ на движущийся зрительный стимул. До этого микроэлектрод был введен в область зоны МТ, и было определено предпочтительное направление движения для клеток этой области, которое отличается от направления движения данного стимула (В). При электрической стимуляции зоны МТ результирующий вектор движения отклоняется в сторону. Эти данные дают основания предполагать, что зрительные движения вычисляются как суммарный вектор нескольких предпочтительных направлений в зоне МТ.
Каким же образом рассчитывается движение в зоне МТ? Колонки в этой зоне представляют собой кластеры клеток, одинаково реагирующих на сходное направление движения вдоль ретинотопической карты. Таким образом, движение объекта вдоль сетчатки должно активировать те колонки, которые имеют ориентацию, совпадающую с направлением движения объекта. Но движущиеся зрительные объекты активируют не только одну подобную колонку. Чаще всего они имеют замысловатую траекторию движения, вызывая активацию большого набора нейронов, чувствительных к различным направлениями движения и к разным его углам. Стало быть, для оценки такого движения и расчета усредненного вектора движения потребуются нейрональные вычисления. Нейрональные вычисления, осуществляемые колонками, чувствительными к направлению движения объекта, изучались Ньюсомом с коллегами, с использованием электрической микростимуляции для управления движения глазом у тренированных обезьян. При помощи микроэлектрода регистрировалось предпочтительное направление движения для данной колонки клеток, а затем этот электрод использовался для инъекции небольшого по величине тока для активирования данной колонки в то время, когда осуществлялось следящее движение глазом (рис. 6). После этого проводили сравнение следящего движения глаза без стимуляции и при стимуляции МТ, для того чтобы узнать, каким образом происходит сложение двух различных направлений движения в пределах зрительного поля. В контроле глаз строго следует за объектом (рис.6А). При стимуляции электрическим током колонки МТ, предпочтения относительно движения которой отличались от направления движения объекта, результирующее движение глаза было средним между двумя этими направлениями (рис. 6В). Вывод: траектория движения используется для расчета среднего вектора для активированных колонок, отслеживающих направление движения. Привлекательной особенностью таких экспериментов является то, что поведение обезьяны (движение глаз) представляет собой тип анализа, выполняемый в высших корковых центрах Уже давно предполагалось, что подобный расчет среднего вектора используется при выполнении резкого отрывистого движения глаз.
Литература
1. Komatsu, H. 1998. Mechanisms of central color vision. Curr. Opin. Neurobiol. 8: 503-508.
2. Merigan, W. H., and Maunsell, J. H. R. 1993. How parallel are the primate visual pathways? Anna. Rev. Neurosci. 16: 369-402.
3. Newsome, W. T., and Wurlz, R. H. 1988. Probing visual cortical function with discrete chemical lesions. Trends Neunsci. 11: 394-400.
4. Tootell, R. В., Dale, A. M., Sereno, M. I., and Malach, R. 1996. New images from human visual cortex. Trends Neurosci. 19: 481-489.
5. Zeki, S. 1990. Colour vision and functional specialisation in the visual cortex. Disc. Neurosci. 6: 1-64.