Реферат: Механизмы дыхания растений
1. Дыхание. Определение. Уравнение. Значение дыхания в жизни растительного организма. Специфика дыхания у растений
Образующиеся в ходе фотосинтеза сахара и другие органические соединения используются клетками растительного организма в качестве питательных веществ. Клетки незеленых частей и все клетки растения в темноте питаются веществами углеводной природы гетеротрофно и в этом принципиально не отличаются от животных клеток. Важнейшим этапом питания органическими веществами на клеточном уровне является процесс дыхания.
Клеточное дыхание — это окислительный, с участием кислорода распад органических питательных веществ, сопровождающийся образованием химически активных метаболитов и освобождением энергии, которые используются клетками для процессов жизнедеятельности.
Научные основы учения о роли кислорода в дыхании были заложены трудами A. Л. Лавуазье. В 1774 г. кислород независимо открыли Пристли и Шееле, а Лавуазье дал название этому элементу. Изучая одновременно процесс дыхания животных и горение, Лавуазье в 1773 — 1783 гг. пришел к выводу. что при дыхании, как и при горении, поглощается 02 и образуется С02, причем в том и другом случаях выделяется теплота. На основании своих опытов он заключил, что процесс горения состоит в присоединении кислорода к субстрату и что дыхание есть медленно текущее горение питательных веществ в живом организме.
Я. Ингенхауз в 1778—1780 гг. показал, что зеленые растения в темноте, а незеленые части растений и в темноте, и на свету поглощают кислород и выделяют С02 так же, как животные. Основателем учения о дыхании растений считают Н. Т. Соссюра. В 1797—1804 гг., впервые широко использовав количественный анализ, он установил, что в темноте растения поглощают столько же кислорода, сколько выделяется С02, т.е. соотношение С02/02, как правило, равно 1. При этом одновременно с С02 образуется и вода. Мнение Соссюра о том. что описанный им газообмен у растений является процессом дыхания и что этот процесс обеспечивает растительный организм энергией, долгое время не признавалось. Утверждалось, что в ночное время растения выделяют тот С02, который не был использован при фотосинтезе, и что этот С02 не имеет отношения к дыханию.
Однако постепенно накапливалось все больше данных о том, что дыхание животных и растений протекает однотипно, несмотря на отсутствие у растений специальных дыхательных органов, причем основным субстратом дыхания служат сахара. И. П. Бородин (1876) в серии точных опытов установил, что интенсивность дыхания листоносных побегов в темноте в первую очередь зависит от количества углеводов, накопленных ими на свету.
Во второй половине XIX в. в результате изучения дыхания у растительных и животных объектов общее уравнение этого процесса приняло следующий вид:
С6Н12О6 + 602 ► 6С02 + 6Н20 + энергия (2875 кДж/моль)
ЗНАЧЕНИЕ ДЫХАНИЯ В ЖИЗНИ РАСТЕНИЯ
Дыхание — один из центральных процессов обмена веществ растительного организма. Выделяющаяся при дыхании энергия тратится как на процессы роста, так и нa поддержание в активном состоянии уже закончивших рост органов растения. Вместе с тем значение дыхания не ограничивается тем, что это процесс, поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно_восстановительный процесc, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения, которые затем используются в различных метаболических реакциях. К промежуточным соединениям относят органические кислоты и пентозы образующиеся при разных путях дыхательного распада. Таким образом, процесс дыхания — источник многих метаболитов. Несмотря на то что процесс дыхания в суммарном виде противоположен фотосинтезу, в некоторых случаях они могут дополнять друг друга. Оба процесса являются поставщиками как энергетических эквивалентов (АТФ, НАДФ-Н), так и метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется также вода. Эта вода в крайних условиях обезвоживания может быть использована растением и предохранить его от гибели. В некоторых случаях, когда энергия дыхания выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. В этой связи при рассмотрении процесса дыхания надо помнить, что не всегда усиление процесса дыхания является полезным для растительного организма.
2.Основные этапы становления учения о дыхании растений
Если, по Лавуазье, дыхание имеет сходство с процессом горения, то каким же образом органические вещества могут «гореть» при обычной температуре тела организма, да еще в водной среде, (ведь на 70 — 90% масса живых организмов состоит из воды)? Возникло предположение о том, что в живых клетках существуют механизмы, активирующие кислород. Швейцарский химик X. Ф. Шейнбайн, открывший озон, изучал причины быстрого потемнения пораненной поверхности растительных тканей, таких, как ткани яблок, картофеля, плодовых тел грибов. В 1845 г. он выступил со своей теорией окислительных процессов, согласно которой в живых клетках имеются соединения, способные легко окисляться в присутствии 02 и таким образом активировать молекулярный кислород. Если ткань прокипятить, то потемнения не происходит. Следовательно, потемнение тканей — каталитический окислительный процесс. Шейнбайн ошибочно полагал, что активация кислорода — это образование озона.
Исследования, начатые Шейнбайном, продолжил А. Н. Бах, который в 1897 г. разработал перекисную теорию биологического окисления, приложив ее к процессам дыхания. Несколько позже, в том же 1897 г., аналогичные взгляды высказал немецкий исследователь К. Энглер.
Суть перекисной теории биологического окисления Баха заключается в следующем. Молекулярный кислород имеет двойную связь и для того чтобы его активировать, необходимо эту двойную связь расщепить. Легко окисляющееся соединение А взаимодействует с кислородом и, разрывая двойную связь, образует пероксид А02 Таким образом, по мысли Баха, активация кислорода есть образование пероксида. В свою очередь пероксидное соединение, взаимодействуя с соединением В, окисляет его; затем эта реакция повторяется со вторым атомом кислорода и второй молекулой соединения В. Получается полностью восстановленное исходное соединение — акцептор кислорода А и полностью окисленное вещество В:
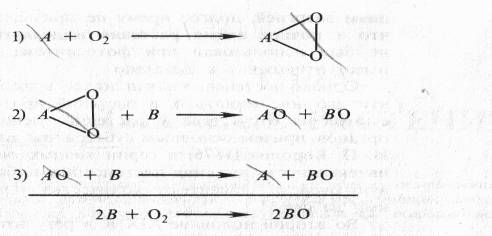
Во второй и третьей реакциях, по Баху, участвует пероксидаза. Соединение А, которое активирует кислород, Бах назвал «оксигеназой», рассматривая его вначале как фермент. В дальнейшем он пришел к заключению, что это те соединения, которые темнеют на воздухе при поранении тканей, что именно такого рода легко окисляющиеся вещества способны присоединять кислород и образовывать пероксиды.
Много позднее, в 1955 г., две группы исследователей — О. Хаяиши с сотр. в Японии и Г. С. Мэзон с сотр. в США, используя современные методы, проанализировали возможность включения кислорода в органические соединения. Применив тяжелый изотоп кислорода, Хаяиши изучил процесс окисления пирокатехина до пирокатеховой кислоты под действием пирокатехазы:
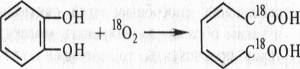
Кислород в пирокатеховой кислоте оказался меченым. Таким образом, в точном эксперименте было доказано, что молекулярный кислород действительно может включаться в органические соединения.
В настоящее время известно, что путь включения кислорода в органические соединения в соответствии с перекисной теорией биологического окисления Баха и Энглера не имеет отношения к дыханию, однако работы этих исследователей сыграли большую роль в изучении химизма дыхания, заложив основы современного понимания механизмов активации кислорода.
Какую же роль играет кислород в процессах дыхания? В 1921 г. немецкий биохимик О. Г. Варбург, изучая влияние ингибиторов на дыхание различных объектов, обнаружил, что поглощение кислорода резко ингибируется оксидом углерода и синильной кислотой, взаимодействующими в клетках с железосодержащим веществом порфириновой природы (с цитохромоксидазой). Английский биохимик Д. Кейлин в 1925 г. окончательно доказал присутствие в клетках цитохромоксидазы, ускоряющей поглощение ими кислорода, и открыл другие цитохромы. Затем цитохромы были обнаружены у всех аэробов и было показано, что у этих организмов на завершающем этапе процесса дыхания осуществляется перенос на кислород электронов и протонов, в результате чего образуется Н20 (или Н202).
Органические соединения могут окисляться и другим способом благодаря отнятию водорода. Еще Бах, сделав упор на перекисную теорию, выдвинул и вторую гипотезу, согласно которой биологическое окисление связано с отнятием от субстрата электронов и протонов. Роль же кислорода состоит в этом случае в регенерации окисленного состояния первичного акцептора водорода. По Баху, последовательность реакций в процессе могла бы выглядеть следующим образом:
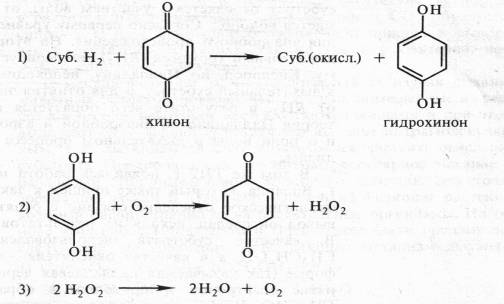
Эта вторая гипотеза Баха в дальнейшем была развита В. И. Палладиным в стройную теорию химизма дыхания. Занимаясь с 1903 по 1916 г. дыхательными пигментами (темнеющими на воздухе веществами тканей), Палладин нашел удачную искусственную модель, которая помогла расшифровать химизм дыхания. Это был известный краситель метиленовый синий. Если в пробирку с раствором метиленового синего поместить несколько прорастающих зародышей пшеницы и выкачать воздух (удалить кислород), то через некоторое время этот краситель полностью обесцветится. Если затем открыть пробирку и встряхнуть содержимое (т. е. дать кислород), то краситель снова синеет. Таким образом, при контакте с кислородом краситель окисляется, приобретая синий цвет, а живая ткань способна восстанавливать краситель, обёсцве чивая его. Причем окисление метиленового синего связано не с присоединением кислорода, а с отнятием водорода. Краситель восстанавливается за счет присоединения электронов и протонов.
На основании этих и других опытов Палладин в 1912 г. в статье «Значение дыхательных пигментов в окислительных процессах растений и животных» представил общую теорию химизма дыхания, разделив основное уравнение дыхания на анаэробную (1) и аэробную (2) части:

где R — это окрашенный дыхательный пигмент, способный отнимать водород от субстрата, a RH2 — бесцветный дыхательный хромоген. На первом, анаэробном, этапе дыхания глюкоза окисляется за счет отнятия водорода, который с помощью фермента (редуктазы) передается на дыхательный пигмент R (активация водорода!). По Палладину, дыхательный субстрат окисляется с участием воды, от которой тоже отнимается водород. Согласно первому уравнению весь С02 дыхания анаэробного происхождения. На втором, аэробном, этапе дыхательный хромоген (RН2) регенерирует в окисленную форму. Кислород, по Палладину, необходим не для включения в дыхательный субстрат, а для отнятия электронов и протонов от RН2, в результате чего образуется вода. В дальнейшем теория Палладина об анаэробной и аэробной фазах дыхания и о роли воды в дыхательном процессе полностью подтвердилась.
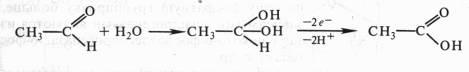
В том же 1912 г. появилась работа немецкого биохимика Г. Виланда, который также пришел к заключению о том, что биологическое окисление связано с отнятием водорода. Этот вывод он сделал, исходя из результатов модельных опытов. В качестве субстрата использовался этиловый спирт СН3СН2ОН, а в качестве окислителя — палладий в губчатой форме (так называемая палладиевая чернь). Происходило отнятие электронов и протонов и образовывался альдегид СН3СНО. Чтобы регенерировать восстановленный палладий и сделать реакцию долгоидущей, Виланд вводил хинон:
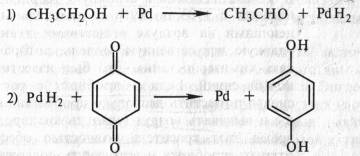
Эта реакция проводилась в небиологической системе и не предусматривала участие кислорода и воды.
Экспериментальные доказательства участия воды в окислении субстрата дыхания и роли 02 как конечного акцептора водорода были получены лишь в 1955 г. в работе Б. Б. Вартапетяна и A. Л. Курсанова. Опыты ставились с этиолированными проростками пшеницы, которым давали 180 в составе 02 или Н20, а через 2 ч анализировали содержание 180 в С02 дыхания
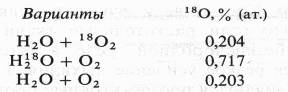
В состав С02 при дыхании тяжелый изотоп кислорода входил из меченой воды, но не из молекулярного кислорода.
Из всех приведенных выше данных следует, что окисление органических веществ в ходе дыхания связано с отнятием водорода и что теория Лавуазье о сходстве дыхания и горения не соответствует действительности.
3.Каталитические системы дыхания
Окисление дыхательных субстратов в ходе дыхания осуществляется с участием ферментов. Ферменты как белковые катализаторы, помимо свойств, присущих неорганическим катализаторам, обладают рядом особенностей: высокой актианостью, высокой специфичностью по отношению к субстратам и высокой лабильностью. Их пространственная организации зависящая от нее активность изменяются под действием внешних и внутренних факторов. Эти свойства обеспечивают возможность тонкой регуляции обмена веществ на уровне ферментов.
Типы окислительно-восстановительных реакций. Существуют четыре способа окисления, и все они связаны с отнятием электронов:
1) непосредственная отдача электронов, например:
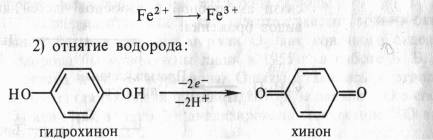
3) присоединение кислорода
![]()
4) образование промежуточного гидратированного соединения с последующим отнятием двух электронов и протонов:
Оксидоредуктазы.
Поскольку окисление одного вещества (донора электронов и протонов) сопряжено с восстановлением другого соединения (их акцептора), ферменты, катализирующие эти реакции, называют оксидоредуктазами. Все они относятся к I классу ферментов:
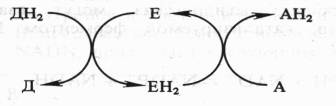
Донор (Д) отдает электроны и протоны, акцептор (А) принимает их, а энзим (Е) осуществляет реакцию переноса. Существуют три группы оксидоредуктаз:
а) анаэробные дегидрогеназы передают электроны различным промежуточным акцепторам, но не кислороду;
б) аэробные дегидрогеназы передают электроны различным акцепторам, в том числе кислороду;
в) оксидазы способны передавать электроны только кислороду.
Анаэробные дегидрогеназы. Это двухкомпонентные ферменты, коферментом которых может быть NAD+ (никотинамидадениндинуклеотид):
При окислении субстрата NAD+ превращается в восстановленную форму NADH, а второй протон субстрата диссоциирует в среду (NADH + Н + ). К анаэробным NAD-зависимым дегидрогеназам относятся такие ферменты, как алкогольдегидрогеназа, лактатдегидрогеназа, малатдегидрогеназа и др. Коферментом анаэробных дегидрогеназ может быть также NADP+ (никотинамидадениндинуклеотидфосфат), содержащий на одну фосфатную группировку больше, чем NAD + . NADP- зависимыми дегидрогеназами являются изоцитратдегидрогеназа, глюкозо-6-фосфатдегидрогеназа, 6-фосфоглюконатдегидрогеназа и др.
Субстратная специфичность фермента зависит от его белковой части. Многие NAD- и NADP-зависимые дегидрогеназы нуждаются в присутствии ионов двухвалентных металлов. Например, алкогольдегидрогеназа содержит ионы цинка.
Окисленные и восстановленные формы коферментов анаэробных дегидрогеназ могут взаимопревращаться в реакции, катализируемой ферментом NAD(Р)-трансгидрогеназой:
NADPH + NAD+ = NADP+ + NADH
Анаэробные дегидрогеназы передают водород, т. е. электроны и протоны, различным промежуточным переносчикам и аэробным дегидрогеназам.
Аэробные дегидрогеназы. Это также двухкомнонентные ферменты, получившие название флавиновых (флавопротеины).
Помимо белков, в их состав входит прочно связанная с ними простетическая группа — рибофлавин (витамин В2).
Различают два кофермента этой группы: флавинмононуклеотид (FMN), или желтый дыхательный фермент Варбурга, и флавинадениндинуклеотид (FAD).
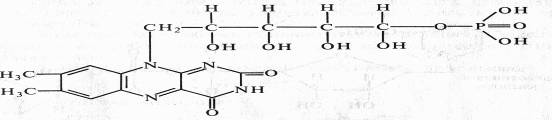
FMN (рибофлавин-5-фосфат) содержит гетероциклическое азотистое основание — диметилизоаллоксазин, спирт рибит (производное рибозы) и фосфат:
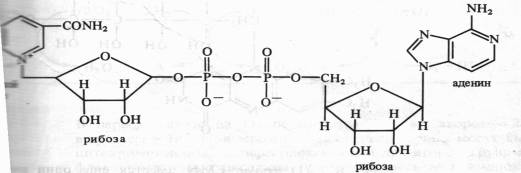
В FAD кроме FMN имеется еще один нуклеотид — аденозинмонофосфата:
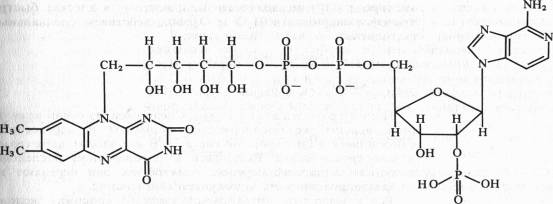
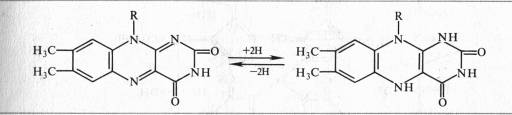
Активной группой в реакции присоединения и отдачи электронов и протонов в FMN и FAD служит изоаллоксазин. Взаимодействие с восстановленным переносчиком, например NADH, происходит следующим образом:
Примером дегидрогеназы, в состав которой входит FAD, является сукцинатдегидрогеназа. Доноры электронов для аэробных дегидрогеназ — анаэробные дегидрогеназы, а акцепторы — хиноны, цитохромы, кислород.
Цитохромная система. Среди оксидаз очень важную роль играют железосодержащие ферменты и переносчики, относящиеся к цитохромной системе. В нее входят цитохромы " и цитохромоксидаза. Включаясь в определенной последовательности в процесс переноса электронов, они передают их от флавопротеинов на молекулярный кислород.
Все компоненты цитохромной системы содержат железопорфириновую простетическую группу.
При переносе электронов цитохромами железо обратимо окисляется и восстанавливается, отдавая или приобретая электрон и изменяя таким образом свою валентность. В дыхательной цепи направление транспорта электронов определяется величиной окислительно-восстановительного потенциала цитохромов.
В этой системе передавать электроны непосредственно на кислород способна только цитохромоксидаза (цит. а + а3). Из всех известных оксидаз она имеет наибольшее сродство к кислороду. Ингибиторами цитохромоксидазы являются СО, цианид, азид. Б растительных митохондриях кроме цитохромоксидазы функционирует оксидаза, не подавляемая цианидом и названная альтернативной оксидазой. Например, в митохондриях початков ароидных активность цианидустойчивой оксидазы в 10 раз превышает активность цитохромоксидазы.
Пероксидаза и каталаза. К пероксидазам относят целую группу ферментов, использующих в качестве окислителя пероксид водорода: классическую пероксидазу, NAD-пероксидазу, NADP-пероксидазу, пероксидазу жирных кислот, глутатионпероксидазу, цитохромпероксидазу и др. Все они работают по следующей схеме, где А — субстраты:
![]()
В последние 2 — 3 десятилетия показана полифункциональность пероксидаз. Помимо пероксидазной, у них имеется оксидазная функция, т. е. способность переносить электроны в отсутствие пероксидного кислорода на молекулярный кислород. Пероксидаза может также функционировать как анаэробная дегидрогеназа, например NADH-дегидрогеназа, передающая электроны от восстановленных пиридиновых нуклеотидов на разные акцепторы.
Пероксид водорода, помимо пероксидазы, расщепляется также каталазой, в результате чего образуется молекулярный кислород. В реакции участвуют две молекулы пероксида, одна из которых функционирует как донор, а другая — как акцептор электронов (см. выше).
Простетической группой пероксидазы и каталазы служит гем, в состав которого входит атом железа.
Оксигеназы. Наряду с оксидазами, которые используют молекулярный кислород как акцептор электронов, в клетках широко представлены оксигеназы, активирующие кислород, в результате чего он может присоединяться к органическим соединениям. Ферменты, внедряющие в субстрат два атома кислорода, называют диоксигеназами, а присоединяющие один атом кислорода — монооксигеназами или гидроксилазами. В качестве доноров электронов оксигеназы используют NAD(P)H, FADH2 и др.
Оксигеназы присутствуют во всех типах клеток. Они участвуют в гидроксилировании многих эндогенных соединений в частности аминокислот, фенолов, стеринов и др., а также в детоксикации чужеродных токсических веществ (ксенобиотиков).
4.Основные пути диссимиляции углерода
Существуют три основных пути диссимиляции углерода. Основной дихотомический путь который включает в себя гликолиз и цикл Кребса. Есть также апотонический путь который включает в себя пентозофосфатный цикл. Путем диссимиляции запасных веществ служит глиоксилатный цикл.
4.1 Дихотомический путь
4.1.1 Гликолиз. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
Гликолиз — процесс анаэробного распада глюкозы, идущий Гликолиз с освобождением энергии, конечным продуктом которого является пировиноградная кислота. Гликолиз — общий начальный этап аэробного дыхания и всех видов брожения. Реакции гликолиза протекают в растворимой части цитоплазмы (цитозоле) и в хлоропластах. В цитозоле гликолитические ферменты, по-видимому, организованы в мультиэнзимные комплексы с участием актиновых филаментов цитоскелета, с которыми гликолитические ферменты обратимо связываются с разной степенью прочности. Такое связывание обеспечивает векторность процесса гликолиза.
Английский биохимик А. Гарден и ученик К. А. Тимирязева Л. А. Иванов в 1905 г. независимо показали, что в процессе спиртового брожения наблюдается связывание неорганического фосфата и превращение его в органическую форму. Гарден установил, что глюкоза подвергается анаэробному распаду только после ее фосфорилирования. Полностью весь процесс гликолиза расшифровали немецкие биохимики Г. Эмбден, О. Ф. Мейергоф и советский биохимик Я. О. Парнас, с именами которых связывают название этого катаболического
Цепь реакций, составляющих суть гликолиза, можно разбить на три этапа (рис. 4.1):
I. Подготовительный этап — фосфорилирование гексозы и ее расщепление на две фосфотриозы.
II. Первое субстратное фосфорилирование, которое начинается с 3-фосфоглицеринового альдегида и кончается 3-фосфоглицериновой кислотой. Окисление альдегида до кислоты связано с освобождением энергии. В этом процессе на каждую фосфотриозу синтезируется одна молекула АТР.
III. Второе субстратное фосфорилирование, при котором 3-фосфоглицериновая кислота за счет внутримолекулярного окисления отдает фосфат с образованием АТР.
Поскольку глюкоза стабильное соединение, на ее активацию необходима затрата энергии, что осуществляется в процессе образования фосфорных эфиров глюкозы в ряде подготовительных реакций. Глюкоза (в пиранозной форме) фосфорилируется АТР с участием гексокиназы, превращаясь в глюкозо-6-фосфат, который изомеризуется в фруктозо-6-фосфат с помощью глюкозофосфатизомеразы. Этот - переход необходим для образования более лабильной фуранозной формы молекулы гексозы. Фруктозо-6-фосфат фосфорилируется вторично фосфофрукгокиназой с использованием еще одной молекулы АТР.
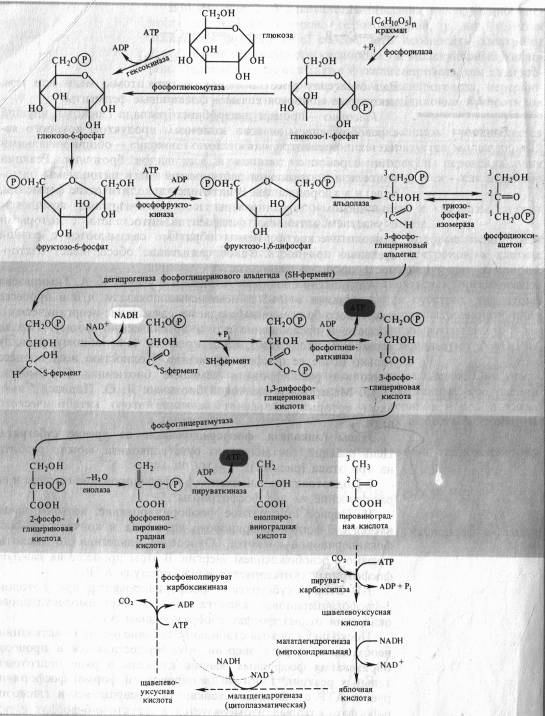
Фруктозо-1,6-дифосфат — лабильная фуранозная форма с симметрично расположенными фосфатными группами. Обе эти группы несут отрицательный заряд, отталкиваясь друг от друга электростатически. Такая структура легко расщепляется аль- долазой на две фосфотриозы. Следовательно, смысл подготовительного этапа состоит в активации молекулы гексозы за счет двойного фосфорилирования и перевода в фуранозную форму с последующим распадом на 3-фосфоглицериновый альдегид (3-ФГА) и фосфодиоксиацетон (ФДА), причем бывший 6-й атом углерода в молекуле глюкозы и фруктозы (фосфорилированный) становится 3-м в 3-ФГК, а 1-й атом углерода фруктозо-1,6-дифосфата остается 1-м углеродом (фосфорилированным) в ФДА (см. рис. 4.1). 3-ФГА и ФДА легко превращаются друг в друга с участием триозофосфатизомеразы. Из-за расщепления молекулы гексозы на две триозы гликолиз иногда называют дихотомическим путем окисления глюкозы.
С 3-ФГА начинается II этап гликолиза — первое субстратное фосфорилирование. Фермент дегидрогеназа фосфоглицеринового альдегида (NAD-зависимый SH-фермент) образует с 3-ФГА фермент-субстратный комплекс, в котором происходит окисление субстрата и передача электронов и протонов на NAD + . В ходе окисления фосфоглицеринового альдегида до фосфоглицериновой кислоты в фермент-субстратном комплексе возникает меркаптанная высокоэнергетическая связь (т. е. связь с очень высокой свободной энергией гидролиза). Далее осуществляется фосфоролиз этой связи, в результате чего SH-фермент отщепляется от субстрата, а к остатку карбоксильной группы субстрата присоединяется неорганический фосфат, причем ацилфосфатная связь сохраняет значительный запас энергии, освободившейся в результате окисления 3-ФГА. Высокоэнергетическая фосфатная группа с помощью фосфоглицераткиназы передается на ADP и образуется АТР. Так как в данном случае высокоэнергетическая ковалентная связь фосфата формируется прямо на окисляемом субстрате, такой процесс получил название субстратного фосфорилирования. Таким образом, в результате II этапа гликолиза образуются АТР и восстановленный NADH.
Последний этап гликолиза — второе субстратное фосфорилирование. З-Фосфоглицериновая кислота с помощью фосфоглицератмутазы превращается в 2-фосфоглицериновую кислоту. Далее фермент енолаза катализирует отщепление молекулы воды от 2-фосфоглицериновой кислоты. Эта реакция сопровождается перераспределением энергии в молекуле, в результате чего образуется фосфоенолпируват — соединение, содержащее высокоэнергетическую фосфатную связь. Таким образом, в этом случае высокоэнергетическая фосфатная связь формируется на основе того фосфата, который имелся в самом субстрате. Этот фосфат при участии пируваткиназы передается на ADP и образуется АТР, а енолпируват самопроизвольно переходит в более стабильную форму — пируват — конечный ппопукт гликолиза.
Энергетический выход гликолиза. При окислении одной молекулы глюкозы образуются две молекулы пировиноградной кислоты. При этом за счет первого и второго субстратного фосфорилирования образуются четыре молекулы АТР. Однако две молекулы АТР тратятся на фосфорилирование гексозы на I этапе гликолиза. Таким образом, чистый выход гликолитического субстратного фосфорилирования составляет две молекулы АТР.
Кроме того, на II этапе гликолиза на каждую из двух молекул фосфотриоз восстанавливается по одной молекуле NADH. Окисление одной молекулы NADH в электронтранспортной цепи митохондрий в присутствии 02 сопряжено с синтезом трех молекул АТР, а в расчете на две триозы (т. е. на одну молекулу глюкозы) - шесть молекул АТР. Таким образом, всего в процессе гликолиза (при условии последующего окисления NADН) образуются восемь молекул АТР. Поскольку свободная энергия гидролиза одной молекулы АТР во внутриклеточных условиях составляет около 41,868 кДж/моль (10 ккал), восемь молекул АТР дают 335 кДж/моль, или 80 ккал. Таков полный энергетический выход гликолиза в аэробных условиях.
Обращение гликолиза. Возможность обращения гликолиза определяется обратимостью действия большинства ферментов, катализирующих его реакции. Однако реакции фосфорилирования глюкозы и фруктозы, а также реакция образования пировиноградной кислоты из фосфоенолпирувата, осуществляемые с помощью киназ, необратимы. На этих участках процесс обращения идет благодаря использованию обходных путей. |Там, где функционируют гексокиназа и фруктокиназа, происходит дефосфорилирование — отщепление фосфатных групп фосфатазами.
Превращение пирувата в фосфоенолпируват также не может осуществиться путем прямого обращения пируваткиназной реакции вследствие большого перепада энергии. Первая реакция обращения гликолиза на этом участке катализируется митохондриальной пируваткарбоксилазой в присутствии АТР и ацетил-СоА (последний выполняет функции активатора).
(Образующаяся щавелевоуксусная кислота (ЩУК), или оксалоацетат, восстанавливается затем в митохондриях до малата при участии NAD-зависимой малатдегидрогеназы (МДГ). Затем малат транспортируется из митохондрий в цитоплазму, где окисляется NAD-зависимой цитоплазматической малатдегидрогеназой снова до ЩУК. Далее под действием ФЕП-карбоксикиназы из оксалоацетата образуется фосфоенолпируват. Фосфорилирование в этой реакции осуществляется за счет АТР (см. рис. 4.1).
Функции гликолиза в клетке. В аэробных условиях гликолиз выполняет ряд функций: 1) осуществляет связь между дыхательными субстратами и циклом Кребса; 2) поставляет на нужды клетки две молекулы АТР и две молекулы NADH при окислении каждой молекулы глюкозы (в условиях аноксии гликолиз, по-видимому, служит основным источником АТР в клетке); 3) производит интермедиа, необходимые для синтетических процессов в клетке (например, фосфоенолпируват, необходимый для образования фенольных соединений и лигнина); 4) в хлоропластах гликолитические реакции обеспечивают прямой путь для синтеза АТР, независимый от поставок NADPH; кроме того, через гликолиз в хлоропластах запасенный крахмал метаболизируется в триозы, которые затем экспортируются из хлоропласта.
Регуляция гликолиза.
Интенсивность гликолиза контролируется в нескольких участках. Вовлечение глюкозы в процесс гликолиза регулируется на уровне Фермента гексокиназы по типу обратной связи: избыток продукта реакции (глюкозо-6-фосфата) аллостерически подавляет деятельность фермента.
Второй участок регуляции скорости гликолиза находится на уровне фосфофруктокиназы. Фермент аллостерически ингибируется высокой концентрацией АТР и активируется неорганическим фосфатом и ADP. Ингибирование АТР предотвращает развитие реакции в обратном направлении при высокой концентрации фруктозо-6-фосфата. Кроме того, фермент подавляется продуктом цикла Кребса — цитратом и через положительную обратную связь активируется собственным продуктом — фруктозо-1,6-дифосфатом (самоусиление).
Высокие концентрации АТР подавляют активность пируваткиназы, снижая сродство фермента к фосфоенолпирувату. Пируваткиназа подавляется также ацетил-СоА.
Наконец, пируВатдегидрогеназный комплекс, участвующий в образовании ацетил-СоА из пирувата, ингибируется высокими концентрациями АТР, а также NADH и собственным продуктом — ацетил-СоА.
4.1.2 Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
В анаэробных условиях пировиноградная кислота (пируват) подвергается дальнейшим превращениям в ходе спиртового, молочнокислого и других видов брожений, при этом NADH используется для восстановления конечных продуктов брожения, регенерируя в окисленную форму. Последнее обстоятельство поддерживает процесс гликолиза, для которого необходим окисленный NAD + . В присутствии достаточного количества кислорода пируват полностью окисляется до С02 и Н20 в дыхательном цикле, получившем название цикла Кребса или цикла ди- и трикарбоновых кислот. Все участки этого процесса локализованы в матриксе или во внутренней мембране митохондрий.
Последовательность реакций в цикле Кребса. Участие органических кислот в дыхании давно привлекало внимание исследователей. Еще в 1910 г. шведский химик Т. Тунберг показал, что в животных тканях содержатся ферменты, способные отнимать водород от некоторых органических кислот (янтарной, яблочной, лимонной). В 1935 г. А. Сент-Дьердьи в Венгрии установил, что добавление к измельченной мышечной ткани небольших количеств янтарной, фумаровой, яблочной или щавелевоуксуснсй кислот резко активирует поглощение тканью кислорода.
Учитывая данные Тунберга и Сент-Дьердьи и исходя из собственных экспериментов по изучению взаимопревращения различных органических кислот и их влияния на дыхание летательной мышцы голубя, английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до С02 через «цикл лимонной кислоты» да счет отнятия водорода. Этот цикл и был назван его именем.
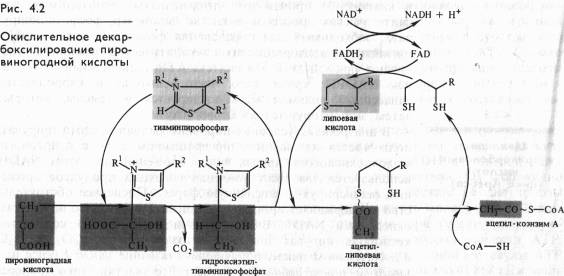
Непосредственно в цикле окисляется ле сам пируват, а его производное — ацетил-СоА. Таким образом, первым этапом на пути окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мультифёрментного комплекса. В состав его входят три фермента и пять коферментов. Коферментами служат тиаминпирофосфат (ТПФ) — фосфорилированное производное витамина Вь липоевая кислота, коэнзим A, FAD и NAD+. Пируват взаимодействует с ТПФ (декарбоксилазой), при этом отщепляется С02 и образуется гидроксиэтильное производное ТПФ (рис. 4.2). Последнее вступает в реакцию с окисленной формой липоевой кислоты. Дисульфидная связь липоевой кислоты разрывается и происходит окислительно-восстановительная реакция: гидроксиэтильная группа, присоединенная к одному атому серы, окисляется в ацетильную (при этом возникает высокоэнергетическая тиоэфирная связь), а другой атом серы липоевой кислоты восстанавливается. Образовавшаяся ацетиллипоевая кислота взаимодействует с коэнзимом А, возникают ацетил- СоА и восстановленная форма липоевой кислоты. Водород липоевой кислоты переносится затем на FAD и далее на NAD + . В результате окислительного декарбоксилирования пирувата образуются ацетил-СоА, С02 и NADH.
Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса. Цикл Кребса начинается с взаимодействия ацетил-СоА с енольной формой щавелевоуксусной кислоты. В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота. Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой. В первой реакции в результате дегидратации лимонной кислоты образуется цис-аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием NAD- или NADP-зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием а-кетоглутаровой кислоты (а-оксоглутаровой кислоты).
а-Кетоглутарат, подобно пирувату, подвергается реакции окислительного декарбоксилирования. а-Кетоглутаратдегидрогеназный мультиэнзимный комплекс сходен с рассмотренным выше пируватдегидрогеназным комплексом. В ходе реакции окислительного декарбоксилирования а-кетоглутарата выделяется С02, образуются NADH и сукцинил-СоА.
Подобно ацетил-СоА, сукцинил-СоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-СоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-CoA может трансформироватся в образование фосфатной связи АТР. При участии сукцинил- СоА-синтетазы из сукцинил-СоА, ADP и Н3Р04 образуются янтарная кислота (сукцинат), АТР, регенерирует молекула СоА. АТР образуется в результате субстратного фосфорилирования.
На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой, коферментом которой является FAD. Фумаровая кислота под действием фумаразы или фумаратгидратазы, присоединяя Н20, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе цикла яблочная кислота с помощью NAD- зависимой малатдегидрогеназы окисляется в щавелевоуксусную. ЩУК, которая самопроизвольно переходит в енольную форму, реагирует с очередной молекулой ацетил-СоА и цикл повторяется снова.
Следует отметить, что большинство реакций цикла обратимы, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзергонические реакции — цитратсинтазная и сукцинил-СоА-синтетазная.
На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул С02, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль Н2О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям — коферментов дегидрогеназ) переносится на кислород .
Выше отмечалось, что цикл Кребса был открыт на животных объектах. Существование его у растений впервые доказал английский исследователь А. Чибнелл (1939).В растительных тканях содержатся все кислоты, участвующие в цикле; обнаружены все ферменты, катализирующие превращение этих кислот; показано, что малонат — ингибитор сункцинатдегидрогеназы — тормозит окисление пирувата и резко снижает поглощение 02 в процессах дыхания у растений. Большинство ферментов цикла Кребса
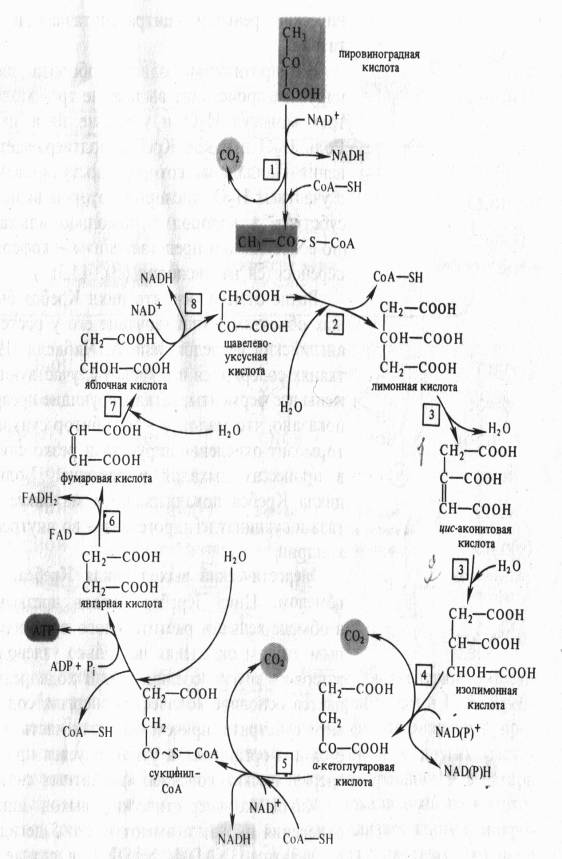
локализовано в матриксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрии.
Энергетический выход цикла Кребса, его связь с азотным обменом. Цикл Кребса. играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии не теряется для организма, а утилизируется при образовании высокоэнергетических конечных фосфатных связей АТР.
Каков же энергетический выход цикла Кребса? В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3NADH, NADPH (в случае изоцитратдегидрогеназы) и FADH2. Окисление каждой молекулы NADH (NADPH) при участии компонентов электронтранспортной цепи митохондрий дает по 3 молекулы АТР, а окисление FADH2 — 2АТР. Таким образом при полном окислении пирувата образуются 14 молекул АТР. Кроме того, 1 молекула АТР синтезируется ; в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекуйы пирувата может образоваться 15 молекул АТР. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТР.
Итак, при окислении глюкозы в процессе дыхания при функционировании гликолиза и цикла Кребса в общей сложности образуются 38 молекул АТР (8 АТР связаны с глико- лизом). Если принять, что энергия третьей сложноэфирнои фосфатной связи АТР равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль).
Регуляция цикла Кребса. Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию.
Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, а-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [NADH]/[NAD + ]. Активность цитратсинтазы тормозится высокой концентрацией АТР и собственным продуктом — цитратом. Изоцитратдегидрогеназа ингибируется NADH и активируется цитратом. а-Кето- глутаратдегидрогеназа подавляется продуктом реакции — сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТР, ADP и восстановленным убихиноном (QH2). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов — высоким уровнем АТР. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена.
Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТР, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТР) продолжается, что поддерживает на низком уровне отношение NADH/NAD+ и снижает уровень АТР. Все это позволяет циклу Кребса функционировать.
4.2 Пентозофосфатный путь. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение. Связь с другими процессами
В клетках растений наряду с гликолизом и циклом Кребса, являющимся главным поставщиком свободной энергии в процессах дыхания, существует и другой важнейший способ катаболизма гексоз — пентоэофосфатный путь (ПФП), в котором участвуют пятиуглеродные сахара (пентозы). Этот путь дыхания известен также как гексозомонофосфатный цикл, пентозный шунт или апотомическое окисление. Окисление глюкозы (глюкозо-6-фосфата) по этому пути связано с отщеплением первого (альдегидного) атома углерода в виде С02 (отсюда и название — апотомический путь).
Пентоэофосфатный путь дыхания открыт в 1935—1938 гг. в результате исследований О. Варбурга, Ф. Диккенса, В. А. Эн- гельгардта и позднее Ф. Липмана. Установлено, что все реакции ПФП протекают в растворимой части цитоплазмы клетс а также в пропластидах и хлоропластах. ПФП дыхания особенно активен в тех клетках и тканях растений, в которых интенсивно идут синтетические процессы, такие, как синтез липидных компонентов мембран, нуклеиновых кислот, клеточных стенок, фенольных соединений.
В ПФП АТР используется только для образования исходного продукта. ПФП, как и цикл Кребса, — циклический процесс, поскольку окисление глюкозы сопровождается регенерацией исходного субстрата ПФП — глюкозо-6-фосфата.
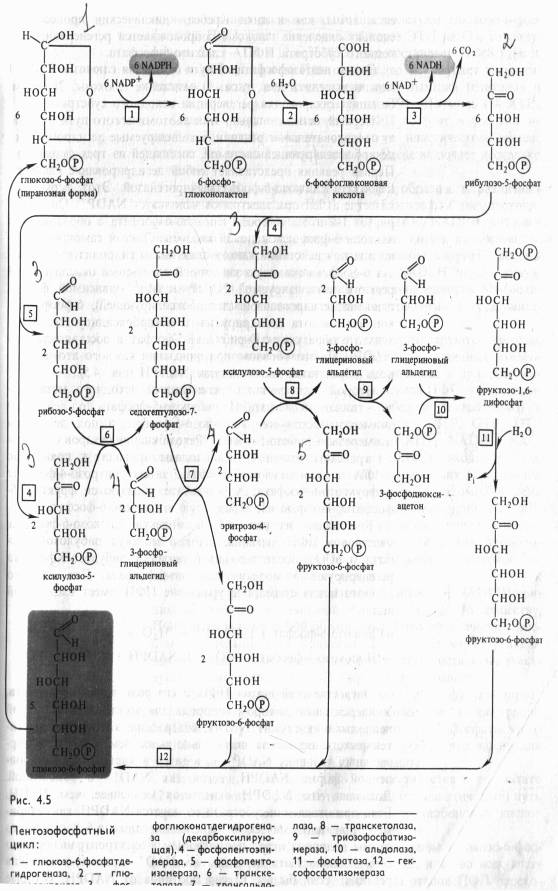
Этапы пентозофосфатного пути окислении глюкозы. В ПФП можно выделить два этапа: 1) окисление глюкозы, 2) рекомбинацию Сахаров для регенерации исходного субстрата.
Первый, окислительный, этап апотомического пути включает последовательные реакции, катализируемые дегидрогеназнодекарбоксилрующей системой, состоящей из трех ферментов. Первая реакция представляет собой Дегидрирование глюкозо- 6-фосфата глюкозо-6-фосфатдегидрогеназой. Этот фермент в качестве акцептора электронов использует NADP + . Он дегидрирует 1-й атом углерода глюкозо-6-фосфата с образованием лактона 6-фосфоглюконовой кислоты. Лактон самопроизвольно или под действием глюконолактоназы гидролизуется, образуя 6-фосфоглюконовую кислоту. В следующей окислительной реакции, катализируемой NADP- и Мп2 +-зависимой фосфо- глюконатдегидрогеназой (декарбоксилирующей), 6-фосфоглюконовая кислота дегидрируется и декарбоксилируется. В результате образуются О-рибулозо-5-фосфат и восстановленный NADPH. Таким образом, при окислении каждого атома углерода образуются две молекулы NADPH (рис. 4.5).
Второй этап связан с регенерацией исходного метаболита — глюкозо-6-фосфата. Из рибулозо-5-фосфата под действием эпимеразы образуется ксилулозо-5-фосфат, а под действием изомеразы — рибозо-5-фосфат. Рекомбинации Сахаров с участием транскетолазы и трансальдолазы приводят к появлению 3-ФГА и седогептулозо-7-фосфата, затем эритрозо-4-фосфата и фруктозо-6-фосфата; в результате образуются фруктозо-6- фосфаты, которые изомеризуются в глюкозо-6-фосфат.
Как видно из рис. 4.5, 6 молекул глюкозо-6-фосфата, участвуя в ПФП дыхания, дают 6 молекул рибулозо-5-фосфата и 6С02, после чего из 6 молекул рибулозо-5-фосфата регенерируют 5 молекул глюкозо-6-фосфата. Для каждого оборота цикла суммарное уравнение ПФП имеет следующий вид:
6 Глюкозо-6-фосфат + 12NADP + + 7Н20->- - 5Глюкозо-6-фосфат + 6С02 + 12NADPH + 12Н+ + Н3Р04
Энергетический выход ПФП и его роль в обмене веществ.
Универсальным донором водорода для электронтранспортной цепи дыхания служит NADH, содержание которого в растительных тканях всегда значительно выше, чем NADPH. В нормальных условиях NADP+ находится в клетках в восстановленной форме NADPH, тогда как NAD+ — в окисленной. Доказано, что NADPH окисляется медленнее, чем NADH. Если при окислении субстрата образуется NADPH, как, например, при апотомическом окислении глюкозо-6-фосфата, то атомы водорода перед поступлением в электронтранспортную цепь должны быть переданы на. NAD+ (трансгидрогеназцая реакция). Если бы все 12 пар протонов от NADPH, которые образуются при полном окислении молекулы глюкозо-6-фосфата по ПФП, были бы переданы через ЭТЦ на 02, то получилось бы 3 АТР х 12 = 36 АТР, что составляет 41,868 кДж х х 36 = 1507 кДж/моль. Практически это не уступает энергетическому выходу дихотомического пути дыхания (гликолиз и цикл Кребса), в котором образуется 1591 кДж/моль (38 АТР,)1
Однако основное назначение ПФП состоит в участии не столько в энергетическом, сколько в пластическом обмене клеток. Это участие в пластическом обмене включает несколько аспектов:
1. NADPH используется главным образом в различных синтетических реакциях.
2.В ходе пентозофосфатного цикла синтезируются пентозы, входящие в состав нуклеиновых кислот и различных нуклеотидов (пиридиновых, флавиновых, адениловых и др.). Для животных и других гетеротрофных организмов ПФП — единственный способ образования пентоз (рибоз и дезоксирибоз) в клетке. Рибозы необходимы для синтеза АТР, GTP, UTP и других нуклеотидов. Коферменты NAD+, NADP+, FAD, коэнзим А — тоже нуклеотиды и в их состав входит рибоза.
3.ПФП имеет большое значение как источник образования углеводов с различным числом углеродных атомов в цепи (от С3 до Су). Эритрозо-4-фосфат, возникающий в ПФП, необходим для синтеза шикимовой кислоты — предшественника многих ароматических соединений, таких, как ароматические аминокислоты, витамины, дубильные и ростовые вещества, лигнин клеточных стенок и др.
4.Компоненты ПФП (рибулозо-1,5-дифосфат, NADPH) принимают участие в темновой фиксации С02. По существу, ПФП представляет собой обращенный фотосинтетический (восстановительный) цикл Кальвина. Только две из 15 реакций цикла Кальвина специфичны для фотосинтеза, остальные участвуют в окислительном ПФП дыхания и гликолизе.
5.В хлоропластах окислительный ПФП функционирует в темноте, предотвращая резкое изменение концентрации NADPH в отсутствие света. Кроме того, триозофосфаты этого цикла в хлоропластах превращаются в 3-ФГК, что важно для поддержания в них уровня АТР в темноте.
Окисление глюкозы по ПФП осуществляется в результате 12 реакций, тогда как в дихотомический (гликолитический) путь через ПВК и далее через цикл ди- и трикарбоновых кислот включаются более 30 различных реакций.
Не следует, однако, думать, что окисление глюкозо-6-фосфата по схеме, представленной на рис. 4.5, идет во всех клетках до конца. Очень часто на одном из этапов ПФП перехо дит в гликолитический. Таким этапом может быть, в частности, транскетолазная реакция (рис. 4.5, реакция 8), в результате которой ксилулозо-5-фосфат и эритрозо-4-фосфат превращаются во фруктозо-6-фосфат и 3-ФГА — субстраты, общие для гликолиза и ПФП.
4.3 Глиоксилатный цикл. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
Этот цикл в 1957 г. был впервые описан у бактерий и плесневых грибов Г. Л. Корнбергом и Г. А. Кребсом. Затем оказалось; что он активно функционирует в про- растающих семенах масличных растений и в других растительных объектах, где запасные жиры превращаются в сахара (глюконеогенез). Глиоксилатный цикл локализован не в митохондриях, как цикл Кребса, а в специализированных микротелах — глиоксисомах. В клетках животных этот цикл отсутствует.
В глиоксилатном цикле из ЩУК и ацетил-СоА синтезируется лимонная кислота, образуются цис-аконитовая и изолимонная (изоцитрат), как и в цикле Кребса . Затем изолимонная кислота под действием изоцитрат-лиазы распадается на глиоксиловую и янтарную кислоты. Глиоксилат с участием малатсинтазы взаимодействует со второй молекулой ацетил-СоА, в результате чего синтезируется яблочная кислота, которая окисляется до ЩУК.
Таким образом, в отличие от цикла Кребса в глиоксилатном цикле в каждом обороте участвует не одна, а две молекулы ацетил-СоА и этот активированный ацетил используется не для окисления, а для синтеза янтарной кислоты. Янтарная кислота выходит из глиоксисом, превращается в ЩУК и участвует, в глюконеогенезе (обращенном гликолизе) и других процессах биосинтеза. Глиоксилатный цикл позволяет утилизировать запасные жиры, при распаде которых образуются молекулы ацетил-СоА.
Регуляция ПФП и глиоксилатного цикла. Пентозофосфатный путь окисления регулируется концентрацией NADP + , так как содержит две NADP-зависимые дегидрогеназы (глюкозо-6- фосфатдегидрогеназу и 6-фосфоглюконатдегидрогеназу). Он регулируется также уровнем синтезов в клетке, потребляющих NADPH (например, синтезом аминокислот и белков). Их высокий уровень приводит к увеличению содержания окисленного NADP + , что стимулирует ПФП.
В регуляции соотношения между ПФП и гликолизом принимает участие ряд интермедиатов: неорганический фосфат, 6-фосфоглюконовая кислота, эритрозо-4-фосфат. Недостаток неорганического фосфата подавляет гликолиз и активирует ПФП. 6-Фосфоглюконовая кислота служит ингибитором гли- колитического фермента фосфофруктокиназы (глюкозофосфатизомеразы), что способствует функционированию ПФП. Эритрозо-4-фосфат, являясь субстратом транскетолазной и трансальдолазной реакций, может тормозить активность ферментов гликолиза и тем самым переключать превращения углеводов с гликолитического на пентозофосфатный путь.
Активность глиоксилатного цикла снижается при повышении концентрации оксалоацетата, который ингибирует сукцинатдегидрогеназу ЦТК. Другой ингибитор цикла — фосфоенолпируват подавляет активность изоцитратлиазы.
5. Цепь переноса водорода и электрона (дыхательная цепь). Комплексы переноса электронов. Окислительное фосфорилирование. Хемиосмотическая теория окисления и фосфорилирования. Механизмы сопряжения процесса транспорта электронов с образованием АТФ
Цикл Кребса, глиоксилатный и пентозофосфатный пути функционируют только в условиях достаточного количества кислорода. В то же время 02 непосредственно не участвует в реакциях этих циклов. Точно так же в перечисленных циклах не синтезируется АТР (за исключением АТР, образующегося в цикле Кребса в результате субстратного фосфорилирования на уровне сукцинил-СоА).
Кислород необходим для заключительного этапа дыхательного процесса, связанного с окислением восстановленных коферментов NADH и FADH2 в дыхательной электронтранспортной цепи (ЭТЦ) митохондрий. С переносом электронов по ЭТЦ сопряжен и синтез АТР.
Дыхательная ЭТЦ, локализованная во внутренней мембране митохондрий, служит для передачи электронов от восстановленных субстратов на кислород, что сопровождается трансмембранным переносом ионов Н + . Таким образом, ЭТЦ митохондрий (как и тилакоидов) выполняет функцию окислительно-восстановительнои Н -помпы. ,
Б. Чане и др. (США) в 50-х годах, используя значения окислительно-восстановительных потенциалов известных в то время переносчиков е~, спектрофотометрические данные о временной последовательности их восстановления и результаты ингибиторного анализа, расположили компоненты ЭТЦ митохондрий в следующем порядке:
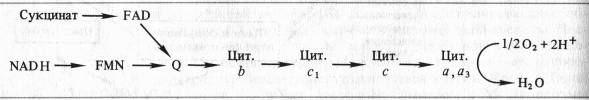
Пара электронов от NADH или сукцината передается по ЭТЦ до кислорода, который, восстанавливаясь и присоединяя два протона, образует воду.
Д. Грин (1961) пришел к выводу, что все переносчики электронов в митохондриальной мембране сгруппированы в четыре комплексу, что было подтверждёно дальнейшими исследованиями.
Согласно современным данным дыхательная цепь митохондрий включает в себя четыре основных мультиэнзимных комплекса и два небольших по молекулярной массе компонента — убихинон и цитохром с
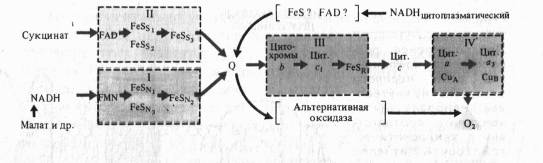
Комплекс I осуществляет перенос электронов от NADH к убихинону Q. Его субстратом служат молекулы вдутримитохондриального NADH, восстанавливающиеся в цикле Кребса. Всостав комплекса входит флавиновая FMN-зависимая NADH: убихи- нон-оксидоредуктаза, содержащая три железосерных центра (FeSN1-3). При встраивании в искусственную фосфолипидную мембрану этот комплекс функционирует как протонная помпа.
Комплекс II катализирует окисление сукцината убихиноном. Эту функцию осуществляют флавиновая (FAD-зависимая) сукцинат : убихинон-оксидоредуктаза, в состав которой также входят три железосерных центра (Fes1_3).
Koмплекс III переносит электроны от восстановленного убихинона к цитохрому с, т. е. функционирует как убихинол: цитохром Т-оксидредуктаза. В своем составе он содержит цитохромы b556 и b560, цитохром с, и железосерный белок Риске. По структуре и функции этот комплекс сходен с цитохромным комплексом b6 — f тилакоидов хлоропластов. В присутствии убихинона комплекс III осуществляет активный трансмембранный перенос протонов.
В терминальном комплексе IV электроны переносятся от цитохрома с к кислороду. _т. е. этот комплекс является цитохромом с: кислород-оксидоредуктазой (цитохромоксидазой). В его состав входят четыре редокс-компонента: цитохромы а и а3 и два атома меди. Цитохром а3 и Сив способны взаимодействовать с 02, на который передаются электроны с цитохрома а — СиА. Транспорт электронов через комплекс IV сопряжен с активным транспортом ионов Н +.
В последние годы в результате изучения пространственного расположения компонентов ЭТЦ во внутренней мембпане митохондрии показано что комплексы I, III и IV пересекают мембрану. На внутренней стороне мембраны, обращенной к матриксу, два электрона и два протона от NADH поступают на флавинмононуклеотид комплекса I.
Электроны передаются на FeS-центры. Пара электронов от FeS-центров захватывается двумя молекулами окисленного убихинона, которые принимают два иона Н + , образуя семихиноны (2QH) и диффундируя к комплексу III. На эти семихиноны поступает еще пара электронов от цитохрома Ь560 комплекса III, что делает возможной реакцию семихинонов с еще двумя протонами из матрикса с образованием 2QH2. Полностью восстановленный убихинон (убихинол) отдает 2е~ цитохрому Ь556 и 2е~ FeSR — цитохрому с,. В результате освобождаются четыре иона Н+, выходящие в межмембранное пространство митохондрии. Окисленные молекулы убихинона вновь диффундируют к комплексу I и готовы принимать от него (или от комплекса II) новые электроны и протоны. Таким образом, цитохромы Ь служат донорами двух электронов для переноса двух дополнительных протонов через липидную фазу мембраны на каждые два электрона, поступающие из комплекса I.
Водорастворимый цитохром с на наружной стороне мембраны, получив 2е~ от FeSR — цитохрома ciy передает их на цитохром а — Сид комплекса IV. Цитохром а3 — Сив, связывая кислород, переносит на него эти электроны, в результате чего с участием двух протонов образуется вода. Как уже отмечалось, цитохромоксидазный комплекс способен также переносить ионы Н+ через митохондриальную мембрану.
Таким образом, из матрикса митохондрии при транспорте каждой пары электронов от NADH к 1/2 02 в трех участках ЭТЦ (комплексы I, III, IV) через мембрану наружу переносятся по крайней мере шесть протонов. Как будет показано далее, именно в этих трех участках окислительные процессы в ЭТЦ сопрягаются с синтезом АТР. Передача 2е~ от сукцината на убихинон в комплексе II не сопровождается трансмембранным переносом протонов. Это приводит к тому, что при использовании сукцината в качестве дыхательного субстрата в ЭТЦ остаются лишь два участка, в которых функционирует протонная помпа.
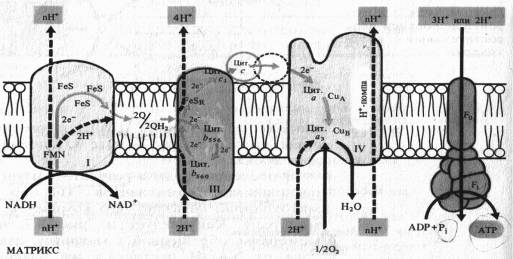
Особенностью растительных митохондрий (ртличающей их от митохондрий животных) является способность окислять экзогенный NADH, т. е. NADH, поступающий из цитоплазмы. Это окисление осуществляется по крайней мере двумя флавиновыми NADH-дегидрогеназами, из которых одна локализована на наружной стороне внутренней мембраны митохондрий, а другая — в их наружной мембране. Первая из них передает электроны в ЭТЦ митохондрий на убихинон, а вторая — на цитохром с. Для функционирования NADH- дегидрогенэзы на наружной стороне внутренней мембраны необходимо присутствие кальция.
Другое существенное отличие растительных митохондрий состоит в том, что во внутренней мембране помимо основного (цитохромного) пути переноса электронов имеется альтернативный путь переноса е~, устойчивый к действию цианида.
Перенос электронов от NADH к молекулярному кислороду через ЭТЦ митохондрий сопровождается потерей свободной энергии. Какова судьба этой энергии? Еще в 1931 г. В. А. Энгельгардт показал, что при аэробном дыхании накапливается АТР. Он первый высказал идею о сопряжении между фосфорилированием ADP и аэробным дыханием. В 1937—1939 гг. биохимики В. А. Белицер в СССР и Г. Калькар в США установили, что при окислении промежуточных продуктов цикла Кребса, в частности янтарной и лимонной кислот, суспензиями животных тканей исчезает неорганический фосфат и образуется АТР. В анаэробных условиях или при подавлении дыхания цианидом такого фосфорилирования не происходит. Процесс фосфорилирования ADP с образованием АТР, сопряженный с переносом электронов по ЭТЦ митохондрий, получил название окислительного фосфорилирования.
Экспериментально установлено, что передача пары электронов от NADH на 02 сопровождается образованием по крайней мерё трех молекул АТР, т. е. коэффициент фосфорилирования Р/О = 3. Такое же значение коэффициента фосфорилирования следует из величин перепадов свободной энергии между различными группами переносчиков. Таких перепадов, достаточных для синтеза молекулы АТР, по крайней мере три: между NADH и FeSN2 в комплексе I («50 кДж), Между убихиноном и цитохромом с1 в комплексе III («13 кДж) и, наконец, между цитохромом а — СиА и 02 (ж84 кДж). Причем если окисляется сукцинат с использованием FAD, то отсутствует первый пункт фосфорилирования и при переносе 2е~ образуются лишь две молекулы АТР.
По поводу механизма окислительного фосфорилирования существуют три теории: химическая, механохимическая (конформационная) и хемиосмотическая.
Химическая и механохимическая шпотезы сопряжения. Согласно химической гипотезе в митохондриях имеются интермёдиаторы белковой природы (X, Y, Z), образующие комплексы с соответствующими восстановленными переносчиками. В результате окисления переносчика в комплексе возникает высокоэнергетическая связь. При распаде комплекса к интермедиатору с высокоэнергетической связью присоединяется неорганический фосфат, который затем передается на ADP:
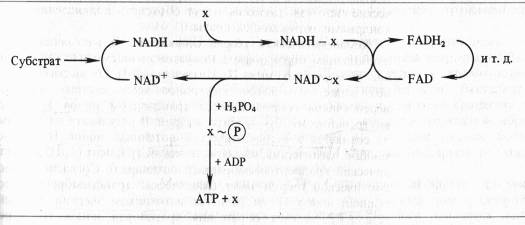
Однако несмотря на упорные поиски, не удалось выделить или как-то иначе доказать реальное существование постулированных высокоэнергетических интермедиаторов типа X ~ P. Гипотеза химического сопряжения не объясняет, почему окислительное фосфорилирование обнаруживается только в препаратах митохондрий с ненарушенными мембранами. И, наконец, с позиций этой гипотезы не находит объяснения способность митохондрий подкислять внешнюю среду и изменять свой объем в зависимости от степени их энергизации.
Способность митохондриальных мембран к конформационным изменениям и связь этих изменений со степенью энергизации митохондрий послужила основой для создания механохимических гипотез образования АТР в ходе окислительного фосфорилирования. Согласно этим гипотезам энергия, высвобождающаяся в процессе переноса электронов, непосредственно используется для перевода белков внутренней мембраны митохондрий в новое, богатое энергией конформационное состояние, приводящее к образованию АТР. Одна из гипотез подобного рода, выдвинутая американским биохимиком П. Д. Бойером (1965), может быть представлена в виде следующей схемы:
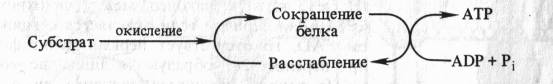
Автор предположил, что запасание энергии происходит путем конформационных изменений ферментов ЭТЦ аналогично тому, как это наблюдается в белках мышц. Актомиозиновый комплекс сокращается, гидролизуя АТР. Если сокращение белкового комплекса достигается за счет другой формы энергии (за счет окисления), то расслабление, возможно, будет сопровождаться синтезом АТР.
Таким образом, согласно механохимическим гипотезам, энергия окисления превращается сначала в механическую энергию, а затем в энергию высокоэнергетической связи АТР. Однако, подобно химической теории сопряжения, механохимические гипотезы также не могут объяснить подкисление митохондриями окружающей среды.
Хемиосмотическая теория сопряжения. В настоящее время наибольшим признанием пользуется хемиосмотическая теория английского биохимика П. Митчелла (1961). Он высказал предположение, что поток электронов через систему молекул-переносчиков сопровождается транспортом ионов Н+ через внутреннюю мембрану митохондрий. В результате на мембране создается электрохимический потенциал ионов Н + , включающий химический, или осмотический, градиент (АрН) и электрический градиент (мембранный потенциал). Согласно хемиосмотической теории электрохимический трансмембранный потенциал ионов Н+ и является источником энергии для синтеза АТР за счет обращения транспорта ионов Н+ через протонный канал мембранной Н + -АТРазы.
Теория Митчелла исходит из того, что переносчики перешнуровывают мембрану, чередуясь таким образом, что в од сторону возможен перенос и электронов, и протонов, а в об ратную — только электронов. В результате ионы Н+ накапливаются на одной стороне мембраны.
Между двумя сторонами внутренней митохондриальной мембраны в результате направленного движения протонов против концентрационного градиента возникает электрохимический потенциа Энергия, запасенная таким образом, используется для синтеза АТР как результат разрядки мембраны при обратном (по концентрационному градиенту) транспорте протонов через АТРазу, которая работает в этом случае как АТР-синтетаза.
За прошедший период хемиосмотическая гипотеза Митчелла получила целый ряд экспериментальных подтверждений. Одним из доказательств роли протонного градиента в образовании АТР при окислительном фосфорилировании может служить разобщающее действие на этот процесс некоторых веществ. Известно, что 2,4-динитрофенол (2,4-ДНФ) подавляет синтез АТР, но стимулирует транспорт электронов (поглощение 02), т. е. разобщает дыхание (окисление) и фосфорилирование. Митчелл предположил, что такое действие 2,4-ДНФ связано с тем, что он переносит протоны через мембрану (т. е. является протонофором) и поэтому разряжает ее. Это предположение полностью подтвердилось. Оказалось, что разные по своей химической природе вещества, разобщающие окисление и фосфорилирование, сходны в том, что, во-первых, они растворимы в липидной фазе мембраны, а, во-вторых, это слабые кислоты, т. е. легко приобретают и теряют протон в зависимости от рН среды. В. П. Скулачев на искусственных фосфолипидных мембранах показал, что чем легче вещество переносит протоны через мембрану, тем сильнее разобщает эти процессы. Другое экспериментальное подтверждение роли протонного градиента в фосфорилировании было получено Митчеллом, который сообщил о синтезе АТР в митохондриях в результате замены щелочной инкубационной среды на кислую (т. е. в условиях искусственно созданного трансмембранного градиента ионов Н+).
В 1973 г. Э. Рэкеру (США) удалось получить липосомы (везикулы из фосфолипидов), в которые была встроена АТРаза, выделенная из митохондрий сердца быка, и хромопротеин галофильной бактерии Halobacterium halobium — бактериородопсин, обусловливающий создание протонного градиента за счет энергии света. Фосфолипиды для реконструкции мембран этих липосом были выделены из растений (соевые бобы). Полученные таким образом гибридные пузырьки на свету осуществляли фосфорилирование.
6.АТФ как основная энергетическая валюта клетки, её структура и функции. Механизмы синтеза АТФ
Процессы обмена вещества включают в себя реакции, идущие с потреблением энергии, и реакции с выделением энергии. В некоторых случаях эти реакции сопряжены. Однако часто реакции, в которых энергия выделяется, отделены в пространстве и во времени от реакций, в которых она потребляется. В процессе эволюции у растительных и животных организмов выработалась возможность хранения энергии в форме соединений, обладающих богатыми энергией-связями. Среди них центральное место занимает аденозинтрифосфат (АТФ). АТФ представляет собой нуклеотидфосфат, состоящий из азотистого основания (аденина), пентозы (рибозы) и трех молекул фосфорной кислоты. Две концевые молекулы фосфорной кислоты образуют макроэргические, богатые энергией связи. В клетке АТФ содержится главным образом в виде комплекса с ионами магния. Аденозинтрифосфат в процессе дыхания образуется из аденозиндифосфата и остатка неорганической фосфорной кислоты (Фн) с использованием энергии, освобождающейся при окислении различных органических веществ: АДФ + ФН --> АТФ + Н2О.При этом энергия окисления органических соединении превращается в энергию фосфорной связи.
В 1939—1940 гг. Ф. Липман установил, что АТФ служит главным нереносчиком энергии в клетке. Особые свойства этого вещества определяются тем, что конечная фосфатная группа легко переноситься с АТФ на другие соединения или отщепляется с выделением энергии, которая может быть использована на физиологические функции. Эта энергия представляет собой разность между свободной энергией АТФ и свободной энергией образующихся продуктов (AG). AG — это изменение свободной энергии системы или количество избыточной энергии, которая освобождается при реорганизации химических связей. Распад АТФ происходит по уравнению AТФ + Н20 = АДФ + ФН, при этом происходит как бы разрядка аккумулятора, при рН 7 выделяется AG = —30,6 кДж. Этот процес катализируется ферментом аденозинтрифосфатазой - (АТФ-аза) Равновесие гидролиза АТФ смещено в сторону завершения peaкции, что и обусловливает большую отрицательную величину свободной энергии гидролиза. Это связано с тем, что при диссоциации. Четырех гидроксильных группировок при рН 7 АТФ имеет четыре отрицательных заряда. Близкое расположение зарядов друг к другу способствует их отталкиванию и, следовательно, отщеплению фосфатных группировок. В результате гидролиза образуются соединения с одноименным зарядом (АДФ3~ и НР04~), которые отживаются друг от друга, что препятствует их соединению. Уникальные свойства АТФ объясняются не только тем, что при ее гидролизе выделяется большое количество энергии, но и тем, что она обладает способностью отдавать концевую фосфатную группу вместе с запасом энергии на другие органические соединения. Энергия, заключенная в макроэргической фосфорной связи, используется на физиологическую деятельность клетки. Вместе с тем по величине свободной энергии гидролиза — 30,6 кДж/моль АТФ занимает промежуточное положение. Благодаря этому система АТФ — АДФ может служить носчиком фосфатных групп от фосфорных соединений с более высокой энергией гидролиза, например фосфоенолпируват (53,6 К/моль), к соединениям с более низкой энергией гидролиза, пример сахарофосфатам (13,8 кДж/моль). Таким образом, система АДФ является как бы промежуточной или сопрягающей.
Механизм синтеза АТР. Сопряжение диффузии протонов назад через внутреннюю мембрану митохондрии с синтезом АТР осуществляется с помощью АТРазного комплекса, получившего название фактора сопряжения F,. На электронно- микроскопических снимках эти факторы выглядят глобулярными образованиями грибовидной формы на внутренней мембране митохондрий, причем их «головки» выступают в матрикс. F1 — водорастворимый белок, состоящий из 9 субъединиц пяти различных типов. Белок представляет собой АТРазу и связан с мембраной через другой белковый комплекс F0, который перешнуровывает мембрану. F0 не проявляет каталитической активности, а служит каналом для транспорта ионов Н+ через мембрану к Fx.
Механизм синтеза АТР в комплексе Fi~ F0 до конца не выяснен. На этот счет имеется ряд гипотез.
Одна из гипотез, объясняющих образование АTP посредством так называемого прямого механизма, была предложена Митчеллом.
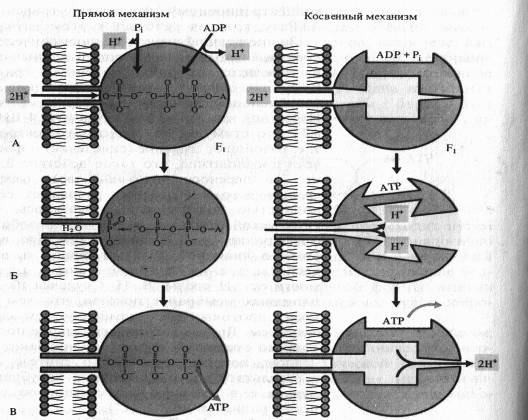
По этой схеме на первом этапе фосфорилирования фосфатный ион и ADP связываются с г компонентом ферментного комплекса (А). Протоны перемещаются через канал в F0-компоненте и соединяются в фосфате с одним из атомов кислорода, который удаляется в виде молекулы воды (Б). Атом кислорода ADP соединяется с атомом фосфора, образуя АТР, после чего молекула АТР отделяется от фермента (В).
Для косвейного механизма возможны различные варианты. ADP и неорганический фосфат присоединяются к активному центру фермента без притока, свободной энергии. Ионы Н + , перемещаясь по протонному каналу по градиенту своего электрохимического потенциала, связываются в определенных участках Fb вызывая конформационныё. изменения фермента (П. Бойер), в результате чего из ADP, и Р( синтезируется АТР. Выход протонов в матрикс сопровождается возвратом АТР-синтетазного комплекса в исходное конформационное состояние и освобождением АТР.
В энергизованном виде F1 функционирует как АТР-синтетаза. При отсутствии сопряжения между электрохимическим потенциалом ионов Н+ и синтезом АТР энергия, освобождающаяся в результате обратного транспорта ионов Н+ в матрикс, может превращаться в теплоту. Иногда это приносит пользу, так как повышение температуры в клетках активирует работу ферментов.
7.Митохондрии как органоиды дыхания. Их структура и функции
Митохондрии — «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4—7 мкм и диаметром 0,5-2 мкм.Число митохондрий в клетке может быть различным, от 500 до 1000. Однако в некоторых организмах (дрожжах) имеется лишь одна гигантская митохондрия. Химический состав митохондрий несколько колеблется. В основном это белковолипоидные органеллы. Содержание белка в них составляет 60—65%. В состав мембран митохондрий входят 50% структурных белков и 50% ферментативных, около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК—1% и ДНК—0,5%. В митохондриях имеется^ не только ДНК, но и вся система синтеза белка, втом числе и рибосомы. Митохондрии окружены двойной мембраной? Толщина мембран составляет 6—10 нм. Между мембранами — перимитохондрлальное пространство, равное 10 нм; оно заполнено жидкостью типа сыворотки. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса.
Внутренняя мембрана дает выросты — кристы, расположенные перпендикулярно продольной оси органеллы и перегораживающие все внутреннее пространство митохондрий на отдельные отсеки. Однако, поскольку выросты-перегородки неполные, между этими отсеками сохраняется связь. Мембраны митохондрий обладают большой прочностью и гибкостью. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов). На внутренней мембране митохондрий расположены грибовидные частицы. Они расположены через правильные промежутки. Каждая митохондрия содержит 104—105 таких грибовидных частиц. Установлено, что в головке грибовидных частиц содержится фермент АТФ-синтетаза, катализирующий образование АТФ аа_счет Энергий, выделяющейся в аэробной фазе дыхания.
Митохондрии способны к движению. Это имеет большое значение в жизни клетки, так как митохондрии передвигаются к тем местам, где идет усиленное потребление энергии. Они могут ассоциировать друг с другом как путем тесного сближения, так и при помощи связующих тяжей. Наблюдаются также контакты митохондрий с эндоплазматической сетью, ядром, хлоропластами. Известно, что митохондрии способны к набуханию, я при потере воды — к сокращению.
В растущих клетках митохондриальный матрикс становится менее плотным, количество крист растет — это коррелирует с увеличением интенсивности дыхания. В процессе дыхания ультраструктура митохондрий меняется. В том случае, если в митохондриях протекает активный процесс преобразования энергии окисления в энергию АТФ, внутренняя часть митохондрий становится более компактной.
Митохондрии имеют свой онтогенез. В меристематических клетках можно наблюдать инициальные частицы, которые представляют собой округлые образования, окруженные двойной мембраной. Диаметр таких инициальных частиц составляет 50 нм. По мере роста клетки инициальные частицы увеличиваются в размере, удлиняются и их внутренняя мембрана образует выросты, перпендикулярные оси митохондрий. Вначале образуются промитохондрии. Они еще не достигают окончательного размера и имеют мало крист. Из промитохондрий образуются митохондрии. Сформировавшиеся митохондрии делятся путем перетяжки или почкованием. Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Сопоставление размеров митохондриальной ДНК с числом и размером митохондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда от митохондрий.
Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б. М. Козо-Полянский высказал мнение, что клетка — это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эта гипотеза имеет много сторонников. Согласно гипотезе симбиогенеза, митохондрии — это в прошлом самостоятельные организмы. По мнению Марголис, это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную содержащую ядро клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение). Сходство обнаруживается и в величине рибосом. Однако доказательств еще недостаточно и окончательного вывода по этому вопросу пока сделать невозможно.
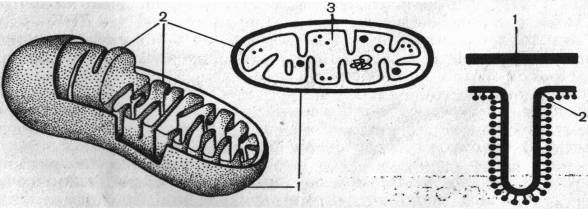
1- наружная мембрана, 2- внутренняя мембрана, 3- матрикс.
8. Генетическая связь дыхания и брожения. Связь дыхания и фотосинтеза. Взаимосвязь дыхания с другими процессами обмена
Соссюр, работая с зелеными растениями в темноте, обнаружил, что они выделяют С02 даже в бескислородной среде. Л. Пастер нашел, что в темноте в отсутствие кислорода в растительных тканях наряду с выделением С02 образуется спирт, т. е. идет спиртовое брожение. Он пришел к выводу, что в растительных тканях, так же как и у бактерий, возможно спиртовое брожение.
Немецкий физиолог Э. Ф. Пфлюгер (1875), изучая дыхание животных объектов, показал, что лягушки, помещенные в среду без кислорода, некоторое время остаются живыми и при этом выделяют С02. Пфлюгер назвал это дыхание интрамолекулярным, т. е. дыханием за счет внутримолекулярного окисления субстрата. Предполагалось, что интрамолекулярное дыхание — начальный этап нормального аэробного дыхания. Эту точку зрения поддержал Б. Пфеффер — немецкий физиолог растений, который распространил ее на растительные организмы. На основе этих работ Пфеффером и Пфлюгером были предложены следующие два уравнения, описывающие механизм дыхания:
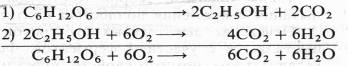
На первом, анаэробном, этапе происходит спиртовое брожение, образуются две молекулы этанола и две молекулы С02. Затем в присутствии кислорода спирт, взаимодействуя с ним, окисляется до С02 и Н20.
С. П. Костычев (1910) пришел к выводу, что это уравнение не соответствует действительности. Он экспериментально доказал, что этанол не может быть промежуточным продуктом нормального аэробного дыхания у растений по двум причинам: во-первых, он ядовит для растений и не может накапливаться, во-вторых, этанол окисляется растительными тканями значительно хуже, чем глюкоза. Костычев предложил свою формулу связи анаэробной и аэробной частей дыхания и различных видов брожения.
В опытах Костычева и его сотрудников (1912—1928) было показано, что если растительные ткани кратковременно выдержать в бескислородной среде, а затем дать кислород, то наблюдается резкое усиление дыхания, т.е. в ходе анаэробной фазы накапливаются промежуточные продукты, которые в присутствии кислорода быстро используются. Ингибиторы, блокирующие брожение, например NaF, блокируют и аэробное дыхание. Ингибиторный анализ (применение ингибиторов специфического действия), выделение и идентификация продуктов окислительного распада глюкозы привели Костычева к выводу о том, что промежуточным продуктом может быть уксусный альдегид. Благодаря работам немецкого биохимика К. Нейберга, Костычева и других стало очевидным, что дыхание и все виды брожения связаны между собой через пировиноградную кислоту (ПВК):
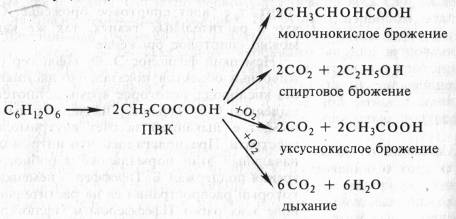
Таким образом, теория Костычева о генетической связи дыхания и брожения полностью подтвердилась.
Взаимосвязь дыхания с другими процессами обмена.
Дыхание тесно связано с другими процессами метаболизма. Необходимо подчеркнуть, что, несмотря на противоположную направленность двух центральных процессов растительного организма —.фотосинтеза и дыхания и на их рассредо- точенность в разных органеллах клетки, между ними существует тесная взаимосвязь. Прежде всего для протекания процесса дыхания органические вещества (субстраты).Таким субстратами в первую очередь являются углеводы, которые у зеленых растений образуются в процессе фотосинтеза. Вместе с тем превращение веществ в процессе фотосинтеза и дыхания идет через ряд сходных промежуточных продуктов. Особенно много сходного в превращениях между фотосинтетическим циклом Кальвина и реакциями пентозофосфатного пути дыхательного обмена. Как в том, так и в другом случае происходят взаимны превращения Сахаров с разной длиной углеродной цепочки (3, 4, 5, 6 и 7 углеродных атомов). По-видимому, несмотря на различное распределение (компартментацию) этих метаболитов в клетке, между ними существует обмен. Иначе го вор я, промежуточные. продукты дыхания могут быть использованы в процессе фотосинтеза. Одновременно возможен и обратный процесс. Много общего в энергетике фотосинтеза и дыхания в процессах фотосинтетического и окислительного фосфорилирования. Между этими двумя процессами возможен обмен энергетическими эквивалентами. АТФ, образовавшаяся на свету при фотосинтетическом фосфорилировании, может служить основным источником энергии для различных биосинтетических процессов, заменяя АТФ, образовавшуюся в процессе дыхания. С другой стороны, АТФ и НАДФ-Н, образовавшиеся в процессе дыхания, могут быть использованы для реакций цикла Кальвина. Имеются наблюдения, что на свету основными органеллами, поставляющими АТФ, являются хлоропласты.
Многие промежуточные продукты процесса дыхания являются основой биосинтеза важнейших соединений. Уже на протяжении первой, анаэробной фазы дыхания (гликолиз) триозофосфат, преобразуясь в глицерин, может служить источником для синтеза жиров. Пировиноградная кислота путем аминирования может дать аланин. Не менее важное значение имеют и промежуточные продукты цикла Кребса. Например, а-кетоглютаровая и щавелевоуксусная кислоты в процессе аминирования дают аминокислоты — глутаминовую и аспарагиновую. Благодаря реакции переаминиро- вания эти кислоты могут быть источником аминогруппы для других аминокислот и, таким образом, являться важнейшими промежуточными продуктами для синтеза как белка, так и пуриновых и пиримидиновых азотистых оснований. Янтарная кислота, образовавшаяся в цикле Кребса, дает основу для образования порфиринового ядра хлорофилла. Ацетил-КоА служит основой для образования жирных кислот. Поскольку имеется ряд реакций и процессов, благодаря которым отдельные компоненты извлекаются из цикла Кребса, должны быть и обратные процессы, поставляющие их в цикл. Если бы этого не было, скорость превращения в аэробной фазе дыхания заметно бы снизилось. Такими реакциями является окислительное дезаминирование аминокислот, приводящее к образованию органических кислот. Имеет значение также реакция карбоксилирования пировиноградной кислоты или ее фосфорилированой формы, в результате чего образуется щавелевоуксусная кислота. Основной процесс, при котором образуются пентозы в растении,— то пентозофосфатный путь дыхательного обмена. Пентозы входят в состав нуклеотидов, нуклеиновых кислот и ряда коферментов, в том числе таких важных, как никотинамидные (НАД и НАДФ), флавиновые (ФМН, ФАД). Пентозофосфатный путь дыхания является также источником образования эритрозо-4-фосфата. Эритрозофосфат, взаимодействуя с фосфоенолпируватом, образует шикимовую кислоту. Шикимовая кислота — материал для образования ряда ароматических аминокислот, например триптофана, а из трипсина образуется один из главных гормонов роста растений — ауксин (Р-индолилуксусная кислота).
Рассмотренные связи дыхания и других процессов метаболизма растения не являются постоянными, раз навсегда данными. Они возникают и нарушаются под влиянием как внутренних особенностей растения, так и внешних условий. При неблагоприятных условиях эти нарушения могут быть значительными и даже летальными.
9.Количественные показатели газообмена
Дыхательный контроль. Возрастание функциональной активности клеток сопровождается усилением дыхания. В значительной степени это достигается благодаря механизму дыхательного контроля, или акцепторного контроля дыхания. Дыхательным .контролем называют зависимость скорости потребления 02 митохондриями от концентрации ADP, который служит акцептором фосфата при окислительном фосфорилировании. В условиях полного сопряжения транспорта электронов по ЭТЦ с синтезом АТР интенсивность дыхательного процесса в митохондриях зависит от концентрации ADP или, точнее, от отношения действующих масс АТР-системы: [ATP]/[ADP] [PJ. Причем неорганический фосфат обычно присутствует в достаточном количестве и не является ограничивающим фактором. В клетке, находящейся в состоянии покоя, это отношение достаточно велико, так как почти весь ADP фосфорилирован. При увеличении функциональной активности клеток АТР расходуется на энергозависимые процессы, в результате чего возрастает концентрация ADP, а это в свою очередь приводит к повышению скорости переноса электронов и интенсивности окислительного фосфорилирования. Важно подчеркнуть, что в данном случае уровень ADP регулирует интенсивность транспорта электронов и окислительное фосфорилирование не как аллостерический фактор, а как субстрат фосфорилирования.
Более полно состояние адениннуклеотидной системы выражается отношением, получившим название энергетического заряда:
[АТР] + у2 [ADP] [АТР] + [ADP] + [AMP] '
который характеризует меру заполнения всей адениннуклеотидной системы высокоэнергетическими фосфатными группами.
10.Регуляция процесса дыхания. Зависимость дыхания от внутренних факторов
Эффект Пастера. Уровень 02 в тканях влияет не только на интенсивность дыхания, но определяет и величину расходования дыхательных субстратов, на что впервые обратил внимание J1. Пастер. В его опытах с дрожжами в присутствии 02 снижались распад глюкозы и интенсивность брожения (уменьшалось количество спирта и выделяемого С02), но одновременно наблюдался интенсивный рост биомассы дрожжей вследствие усиления использования Сахаров на синтетические процессы. Торможение распада Сахаров и более эффективное их использование в присутствии кислорода получило название «эффекта Пастера».
Механизм эффекта Пастера состоит в том, что в присутствии 02 интенсивно идущий процесс окислительного фосфорилирования конкурентно уменьшает количество молекул ADP, вступающих в гликолиз (на нужды субстратного фосфорилирования). По этой причине, а также из-за тормозящего действия АТР (синтез которого резко возрастает в аэробных условиях) на фосфофруктокиназу, скорость процессов гликолиза в присутствии 02 снижается. Избыток АТР может способствовать и ресинтезу глюкозы из части молекул пирувата, образующегося в ходе гликолиза. Без кислорода не функционируют цикл Кребса и ПФП и, следовательно, клетки не получают многих промежуточных соединений, необходимых для синтеза клеточных структур. В присутствии 02 все эти циклы работают. Увеличение концентрации молекул АТР в условиях аэробиоза также способствует синтетическим процессам.
11.Зависимость процесса дыхания от факторов внешней среды
Концентрация 02. Процесс дыхания связан с непрерывным потреблением кислорода клетками и тканями растений. В то же время окислительные превращения субстратов при дыхании включают не только аэробные, но и анаэробные процессы (гликолиз, брожение). Поэтому интересен тот факт, что при снижении парциального давления 02 с 21 до 9% интенсивность дыхания тканей растений меняется незначительно. При 5% 02 у молодых растений начинает несколько снижаться поглощение 02 и в меньшей степени изменяется выделение С02. Как отмечалось, одна из важнейших конечных оксидаз аэробного дыхания — цитохромоксидаза — обладает высоким сродством к кислороду и может функционировать при его низких парциальных давлениях.
Эти факты можно объяснить как тем, что в эволюции дыхательные системы растений формировались в низкокислородных условиях, так и тем, что возможности функционирования аэробной дыхательной цепи при низких парциальных давлениях 02 — один из важных адаптивных признаков, элемент надежности энергетики растения. Это тем более важно, что газовый состав внутренних тканей органов растений может отличаться от атмосферного. Так, в листовой паренхиме сахарной свеклы содержание 02 меняется в течение суток от 7,1 до 17,4%, а С02 - от 0,9 до 5,1%; в мякоти зрелых яблок содержится 7,5 С02 и 13,9% 02, а плодов лимона — 8,5 С02 и 11,5% 02.
У растений, обитающих в условиях систематического затопления (болотные и др.), существует ряд приспособлений к перенесению недостатка кислорода: развитие аэренхимы, способность использовать кислород нитратов («нитратное» дыхание), различные способы устранения избытка продуктов брожения (этанола, молочной кислоты), а также механизмы их использования на нужды обмена веществ.
Избыток кислорода в тканях растений может возникать лишь локально. В атмосфере чистого кислорода дыхание растений снижается, а при длительном его действии растение погибает. Эффект избытка 02 связан с усилением в клетках свободнорадикальных реакций, повреждением мембран вследствие окисления их липидов и в конечном итоге — с нарушением многих процессов обмена веществ.
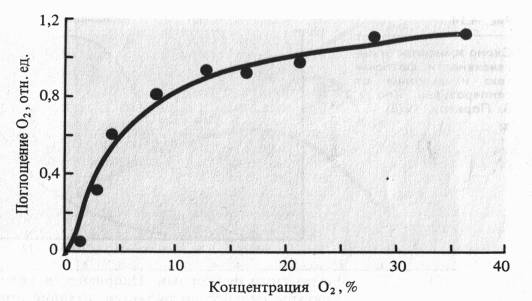
Диоксид углерода. Повышение концентрации С02 как конечного продукта дыхания приводит к снижению интенсивности дыхания. При повышении концентрации С02 тормозятся реакции декарбоксилирования и активность сукцинатдегидрогеназы и, следовательно, уменьшаются выделение С02 и дыхательный коэффициент. При этом наблюдается закисление тканей — ацидоз, что может приводить к вредным последствиям. С02 хорошо растворяется в липидах и таким образом может воздействовать на мембраны. По-видимому, он обладает некоторым наркотическим действием. Предполагается, что С02 способен регулировать метаболизм растений в анаэробных условиях. У листьев ингибирующее действие высоких концентраций С02 на дыхание может быть связано с закрыванием устьиц в этих условиях. Повышенное содержание С02 в тканях семян, покрытых плотной оболочкой, — один из способов поддержания состояния покоя.
Температура. Дыхание, подобно другим ферментативным процессам, зависит от температуры. В определенных температурных границах эта зависимость подчиняется правилу Вант-Гоффа (скорость химических реакций удваивается при повышении температуры на 10 °С). В интервале температур от 0 до 20 °С <210 дыхания равен 2 — 3. При температурах выше 20 °С величина Ql0 может понижаться. Одна из причин этого — уменьшение растворимости 02 в жидкостях при повышении темпепатупы. ЭАсЬект темпепатупы проявляется обычно вместе с другими факторами. Например, в условиях высокой температуры сильнее подавляется дыхание при пониженном содержании 02 или повышенной концентрации С02 и т. д.
Для дыхания каждого вида растений и его органов существуют определенные минимальные (когда дыхание еще возможно), оптимальные и максимальные температуры. В отличие от фотосинтеза дыхание у зимующих растений наблюдается при очень низких температурах (у хвои ели и сосны при — 25 °С). Температурный оптимум его у большинства видов умеренных широт лежит в пределах 35 — 40 °С, т. е. на 5 — 10 °С выше, чем для фотосинтеза. Максимальные температуры (45 — 55 °С) определяются способностью белков к денатурации.
При рассмотрении влияния на дыхание крайних температур важно учитывать продолжительность действия этого неблагоприятного фактора. Так, при кратковременных экспозициях (30 мин) температурный оптимум дыхания листьев гречихи сахалинской может быть очень высок (> 47 °С), тогда как при двухчасовом действии оптимум смещается к 44 °С, а при шестичасовом—до 42 °С.
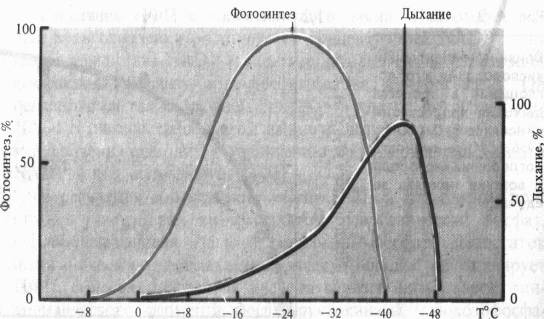
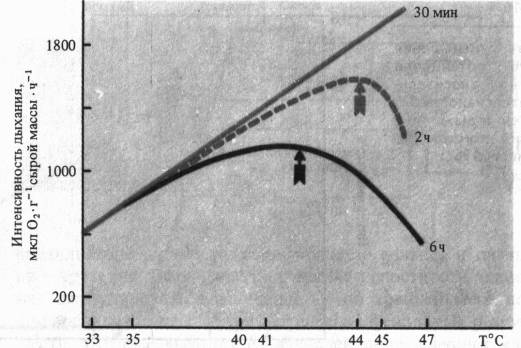
Водный режим. Изменение оводненности тканей растения отражается на интенсивности дыхания. В листьях проростков при быстрой потере воды вначале отмечается усиление дыхания — обычная кратковременная реакция на раздражение. При постепенном снижений оводненности этого не происходит. Продолжающийся водный дефицит вызывает постепенное снижение дыхания, причем более медленное, чем падение интенсивности фотосинтеза. Более раннее прекращение фотосинтетической активности при водном дефиците является следствием закрытия устьиц (уменьшение поступления С02 в лист) и потери тургорного давления в клетках.
Необходимость определенной степени оводненности тканей для процесса дыхания отчетливо проявляется при изучении дыхания семян. Дыхание воздушно-сухих семян (10—11% Н20) ничтожно мало. При повышении влажности семян до 14—15% дыхание возрастает в 4 —5 раз, а при дальнейшем повышении оводненности семян до 30 — 35% дыхание увеличивается в тысячи раз. При этом важную роль играет температура, при которой хранятся семена: при 0— 10°С влияние влажности на интенсивность дыхания значительно меньше, чем при 18 — 25 °С. Резкий подъем дыхания набухших семян сопровождается значительным выделением тепла, что может привести к их перегреву («самовозгоранию») при хранении.
Реакция растительных тканей на потерю воды зависит также от возраста, физиологического состояния, экологической принадлежности организма.
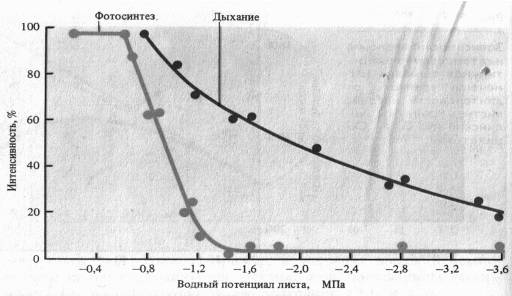
Минеральные вещества. Добавление раствора солей в воду, на которой выращивают проростки, обычно усиливает дыхание корней. Этот эффект получил название «солевого дыхания». Полагают, что солевое дыхание отражает усиление обмена веществ, необходимое для энергетической поддержки активного транспорта ионов корнем. Однако до конца это явление не изучено. Отмечены случаи, когда увеличенная интенсивность дыхания сохранялась после удаления солей. В тканях других органов этот эффект удается получить не всегда.
Свет. Действие света на дыхание зеленых органов растений из-за методических трудностей изучено недостаточно, так как одновременно с дыханием в них осуществляется фотосинтез. Освещенность, при которой интенсивность фотосинтеза равна интенсивности дыхания (по уровню поглощенного и выделенного С От), называется компенсационным пунктом.
При работе с незелеными растениями или с незелеными органами растений возможно непосредственное изучение действия света на дыхание. Дыхание (поглощение 02 и выделение С02) незеленых тканей активируется светом коротковолновой части спектра — ближним ультрафиолетом (380 нм), синими и зелеными лучами (400 — 500 нм) .
В области 380 — 600 нм лежат максимумы поглощения каротиноидов, флавинов и всех цитохромов дыхательной цепи митохондрий. Усиление дыхания может быть связано с прямым действием света на эти компоненты окислительно-восстановительных систем клеток.
Остается нерешенным вопрос о характере взаимодействия процессов фотосинтеза и дыхания в зеленых клетках в ходе фотосинтеза. Обсуждаются две точки зрения об использовании АТР, образующегося при фотофосфорилировании. Согласно одной из них на свету хлоропласта большую часть синтезируемой при фотофосфорилировании АТР используют на синтез углеводов, отдавая на нужды клетки лишь небольшую ее часть. АТР в цитоплазму поставляют митохондрии, которые используют ассимиляты. В темноте хлоропласта сами нуждаются в поступлении энергии из цитоплазмы.
По другой гипотезе АТР фотосинтеза на свету обеспечивает энергией не только фиксацию С02, но и другие энергозависимые процессы в клетке. Окислительное фосфорилирование в митохондриях на свету резко замедляется, а цикл Кребса ускоряется, так как его интермедиаты необходимы для функционирования хлоропластов.
Список использованной литературы
1.В.В. Полевой Физиология растений М.: Высшая школа 1989г.
2.Н.И. Якушкина Физиология растений М.: Просвещение 1993г.
| Биологическая роль, структура и выделение митохондрий из печени крыс. | |
|
Содержание 1. Введение История вопроса Перспективы исследований Биологическая роль митохондрий Ультраструктура митохондрий Общие принципы организации ... То обстоятельство, что ферменты дыхательной цепи составляют более 25 % белка митохондриальных мембран заставляют рассматривать эти ферменты не только как функциональные, но и как ... Так, благодаря наличию в составе митохондрий различных белков, липидов, нуклеиновых кислот и углеводов, соединению их между собой и упорядоченному размещению в пространстве в виде ... |
Раздел: Рефераты по биологии Тип: реферат |
| Развитие, становление и основные аспекты фармации | |
|
РАЗВИТИЕ, СТАНОВЛЕНИЕ И ОСНОВНЫЕ АСПЕКТЫ ФАРМАЦИИ Для ветеринарного провизора необходимы знания, с помощью которых можно контролировать качество ... Серу, содержащуюся в молекулах производных сульфокислот, сульфаниламидных препаратов, метансульфат-ионе, можно обнаружить озолением, окислением (концентрированной азотной кислотой ... Первая группа цветных реакций позволяет обнаружить наличие стероидного цикла в молекуле, например реакция Либермана-Бурхардта, основанная на способности стероидов к дегидратации ... |
Раздел: Рефераты по медицине Тип: книга |
| Общая и неорганическая химия | |
|
Квантово-механическая модель атома. Квантовые числа. Атомные орбитали. Порядок заполнения орбиталей электронами Теория строения атома основана на ... Образуется между наиболее электроотрицательными элементами (фтор, азот, кислород), реже - в некоторых других (хлор, сера).Наиболее прочные водородные связи имеются в воде ... Мембрана, окружающая нормальную клетку крови, проницаема лишь для молекул воды, кислорода, некоторых из растворенных в крови питательных веществ и продуктов клеточной ... |
Раздел: Рефераты по химии Тип: учебное пособие |
| Мембранная энзимология | |
|
... не просто некая пассивная структура, ограничивающая водные компартменты. Уже краткое знакомство с типами ферментов, связанных с мембранами, показывает Примерами таких ферментов могут служить цитохром с-оксидаза, катализирующая перенос электронов через мембрану и транспорт протонов в противоположном направлении, а также ... Большинство работ посвящено связыванию ферментов цикла трикарбоновых кислот и /3-окисления жирных кислот с внутренней митохондриальной мембраной и ферментов гликолиза с ... |
Раздел: Рефераты по биологии Тип: реферат |
| Методы определения концентрации растворённого кислорода в воде | |
|
Пояснительная записка к курсовой работе тема: " Методы определения концентрации растворённого кислорода в воде" Введение Контроль содержания кислорода ... Кислород может быть получен в лаборатории также рядом других методов, из которых наиболее удобны следующие: а) слабое прокаливание КМnO4; б) приливание по каплям раствора КМnO4 к ... Молекулы кислорода диффундируют к поверхности электрода и, сорбируясь на нём, диссоциируют на атомы и ионизируются за счёт электронов электрода, проникая к границе раздела проба ... |
Раздел: Рефераты по химии Тип: курсовая работа |